Autor: William M. Baum
Tradutores: Diego Lima, Lunna Toffano de Abreu e Lima, Karollane Sapucaia, Andresa De Souza, Guilherme Chirinéa
REPENSANDO O REFORÇAMENTO: ALOCAÇÃO, INDUÇÃO E CONTINGÊNCIA
O conceito de reforçamento é pelo menos incompleto e quase certamente incorreto. Uma forma alternativa de organizar nosso conhecimento sobre comportamento pode ser construída em torno de três conceitos: alocação, indução e correlação. Alocação é a medida do comportamento e capta a centralidade da escolha: todo comportamento implica em escolhas e consiste em escolhas. Alocação muda como resultado de indução e correlação. O termo indução abrange fenômenos como comportamentos adjuntivos, interinos e terminais – comportamentos induzidos em uma situação por ocorrência de comida ou outro Evento Filogeneticamente Importante (Phylogeneticly Important Event – PIE) na situação. A indução se assemelha ao controle de estímulos no fato de que não existe relação de um-para-um entre o evento que induz e o comportamento induzido. Se se permitisse que algum controle de estímulo fosse o resultado de filogenia, então a indução e o controle de estímulos seriam idênticos, e um PIE seria semelhante a um estimulo discriminativo. Muitas evidências sustentam a ideia que um PIE induz todas atividades relacionadas ao PIE. Pesquisas também apoiam a ideia de que estímulos correlacionados a PIEs se tornam indutores condicionais relacionados a PIEs. Contingências criam correlações entre atividades “operantes” (como pressão à barra) e PIEs (como comida). Quando uma atividade se torna relacionada a um PIE, ela é induzida pelo PIE bem como outras atividades ligadas a PIEs. Contingencias também restringem possíveis performances. Essas restrições também especificam funções de retorno (feedback), o que explica fenômenos como a maior taxa de respostas em esquemas de razão comparado a esquemas de intervalo. Alocações que incluem muitas atividades operantes são “selecionadas” apenas no sentido em que elas geram uma frequência maior da ocorrência do PIE na situação: contingencia e indução fazem a “seleção”.
Palavras chave: reforçamento, contingencia, correlação, indução, estímulo indutor, indutor, alocação, Evento Filogeneticamente Importante, restabelecimento
_________________________
O objetivo desse artigo é expor um modelo conceitual para compreender o comportamento em relação ao ambiente. Não tentarei explicar todos os fenômenos conhecidos pelos analistas do comportamento; seria impossível. Em vez disso, vou oferecer esse modelo como uma forma de pensar nesses fenômenos. A intenção é substituir o modelo tradicional, que já tem mais de 100 anos, segundo o qual reforçadores supostamente fortalecem a resposta ou a conexão estimulo-resposta, e no qual o condicionamento clássico e condicionamento operante são considerados dois processos distintos. Espera-se que leitores que conhecem esse assunto não encontrem nenhuma novidade aqui, pois os pedaços desse modelo já existiam, e eu simplesmente juntei-os. Ele baseia-se em três conceitos: (1) alocação, que é a medida do comportamento; (2) indução, que é o processo motivador do comportamento; e (3) contingência, que é a relação que compele e conecta eventos comportamentais e ambientais. Como nenhum destes existe em um dado momento, eles necessariamente implicam em uma visão molar do comportamento.
Conforme expliquei em trabalhos anteriores (Baum, 2001; 2002; 2004), as visões molar e molecular não competem entre si, são paradigmas diferentes (Kuhn, 1970). A escolha entre elas não é feita com base em dados, mas baseada em plausabilidade e elegância. Nenhum teste experimental pode escolher entre elas, pois são incomensuráveis – ou seja, diferem ontologicamente. A visão molecular diz a respeito a respostas discretas, estímulos discretos e contiguidade entre estes eventos. Ela oferece estes conceitos para a construção de teorias. Foi projetada para explicar mudanças no comportamento a curto prazo, abruptas, tais como as que acontecem em registros cumulativos (Ferster & Skinner, 1957; Skinner, 1938). Já quando aplicada a fenômenos temporalmente estendidos, como escolha, os resultados são fracos, pois suas teorias e explicações quase sempre lançam mão de construtos hipotéticos para lidar com lacunas temporais, o que faz com que sejam implausíveis e deselegantes (Baum, 1989). A visão molar refere-se a atividades extensas, contextos extensos e relações extensas. Efeitos a curto prazo são tratados como fenômenos localizados, menos extensos (Baum, 2002; 2010; Baum & Davison, 2004). Como o comportamento, por sua própria natureza, necessariamente se desenrola no tempo (Baum, 2004), as teorias e explicações construídas baseando-se na visão molar tendem a ser simples e diretas. Qualquer teoria molar ou molecular específica pode ser invalidada pelo teste experimental, mas ninguém deveria pensar que o paradigma é portanto invalidado; uma nova teoria sempre pode ser inventada dentro do paradigma. O paradigma molar supera o paradigma molecular produzindo teorias e explicações que são mais plausíveis e elegantes.
A Lei do Efeito
E.L. Thorndike (2000/1911), ao propor a lei do efeito, escreveu: “De várias respostas à mesma situação, aquelas que são acompanhadas ou seguidas de perto pela satisfação da vontade do animal, sendo as outras coisas mantidas iguais, são mais firmemente conectadas com a situação, de maneira que, quando esta ocorrer novamente, terão maior probabilidade de ocorrer novamente…” (p. 244).
Desde o início, a lei foi bastante criticada. J.B. Watson (1930), ridicularizando a teoria de Thorndike, escreveu: “A maioria dos psicólogos… acreditam que a formação de hábitos é implantada por fadas bondosas. Por exemplo, Thorndike fala em prazer carimbando o movimento bem sucedido e desprazer apagando os movimentos mal sucedidos.” (p. 206).
Assim, já em 1930, Watson era cético com relação à ideia de que consequências satisfatórias poderiam fortalecer respostas. Ainda assim, a teoria continuou apesar de criticas ocasionais (e.g. Baum, 1973; Staddon, 1973).
A teoria de Thorndike se tornou a base da teoria do reforçamento de B. F. Skinner. No entanto, Skinner fez uma distinção entre observação e teoria. Em seu trabalho “Teorias de aprendizagem são necessárias?” Skinner (1950) escreveu “… a Lei do Efeito não é nenhuma teoria. Ela simplesmente especifica um procedimento para alterar a probabilidade de uma dada resposta.”
Então, Skinner defendeu que a Lei do Efeito se referia à observação de que quando, por exemplo, alimento torna-se contingente à pressão à barra, a pressão a barra aumenta em frequência. No entanto, assim como Thorndike, ele tinha uma teoria de como isso aconteceu. Em seu trabalho de 1948 “Superstição no Pombo”, Skinner reafirmou a teoria de Thorndike com uma nova linguagem: “Dizer que reforçamento é contingente a uma resposta pode não significar nada além do que ele segue a resposta… provavelmente o condicionamento acontece apenas por causa da relação temporal, expressa em termos da ordem e proximidade entre resposta e reforçamento.”
Se substituirmos “satisfação” por “reforçamento” e “acompanhado ou seguido de perto” por “ordem e proximidade” retornaríamos à teoria de Thorndike acima.
Skinner persistiu em sua visão que ordem e proximidade entre resposta e reforçador são a base do reforçamento; isso apareceu novamente em Ciência e Comportamento Humano (Skinner, 1953): “No que se refere ao organismo, a única propriedade importante da contingência é temporal. O reforçador apenas segue a resposta… Devemos supor que a apresentação de um reforçador sempre reforça algo, já que necessariamente coincide com algum comportamento.” (p.85).
Da presente perspectiva, sabemos que a observação sobre contingência de Skinner estava correta. No entanto, sua teoria sobre ordem e proximidade estava incorreta, pois um “reforçador” não “reforça” aquilo com o qual coincide. Isso não acontece no dia-a-dia; se eu por acaso estiver assistindo televisão quando o entregador de pizzas chegar, estarei eu inclinado a assistir mais televisão? (Embora, como veremos, a pizza importe, à medida que agrega valor a, digamos, assistir um jogo de futebol na televisão.) Também não acontece em laboratório, como Staddon (1977), entre outros, mostrou.
Quando Staddon e Simmelhag (1971) repetiram o experimento de “superstição” de Skinner, eles observaram que muitas atividades diferentes ocorriam durante os intervalos entre as apresentações de alimento. Significativamente, as atividades muitas vezes não eram seguidas de perto por alimento; algumas apareciam depois da apresentação do alimento e desapareciam assim que ele era apresentado. A Figura 1 mostra alguns dados apresentados por Staddon (1977). O gráfico à esquerda mostra os efeitos de apresentar alimento a cada 12s para um pombo faminto. As atividades de se aproximar da parede da janela da câmara e de bater as asas aumentou e depois desapareceu, minando qualquer noção de que foram acidentalmente reforçadas. O gráfico à direita mostra o efeito de apresentar alimento a um rato faminto a cada 30s. No começo do intervalo o rato bebe e corre, mas estas atividades desapareceram antes que pudessem ser seguidas por alimento. Pesquisas subsequentes confirmaram estas observações muitas vezes, contradizendo a noção de fortalecimento por contiguidade acidental (e.g. Palya & Zacny, 1980; Reid, Bacha & Morán, 1993; Roper, 1978).

Fig 1. Comportamento induzido por alimento periódico. Esquerda: atividades realizadas por pombo apresentado a alimento a cada 12s. Direita: atividades realizadas por rato apresentado a alimentado à cada 30s. Atividades que desapareceram antes do alimento ser entregue não puderam ser reforçados. Reimpresso de Staddon (1977).
Diante de resultados como os da Figura 1, alguém teorizando com base no paradigma molecular tem pelo menos duas opções. Primeiramente, a contiguidade entre respostas e o alimento pode ser esticada para sugerir que as respostas são reforçadas de forma fraca diante de atraso. Segundo, a contiguidade alimento-resposta pode ser exercida baseando-se na ideia de que o alimento pode eliciar respostas. Por exemplo Killeen (1994) teorizou que cada entrega de alimento eliciava excitação, um construto hipotético. Ele propôs que a excitação saltava seguindo o alimento e então se dissipava depois de tempo sem alimento. Nesse caso, o desenvolvimento da excitação supostamente causava as respostas da Figura 1. Como veremos mais a frente, o paradigma molar evita a entidade hipotética ao basear-se por sua vez na relação estendida de indução.
Para alguns pesquisadores, a inadequação do reforço baseado em contiguidade pode não ser novidade (Baum, 1973). Uma resposta para essa inadequação tem sido ampliar o conceito de reforçamento tornando-o sinônimo de optimização ou adaptação (e.g. Rachlin, Battalio, Kagel & Green, 1981; Rachlind & Burkhard, 1978). No entanto, essa redefinição requer questionamentos sobre o mecanismo. Assim como Herrnstein (1970) escreveu: “Retroceder ao senso comum e concluir que os animais são adaptativos, i.e., fazem o que mais lhe traz beneficio, é uma tentação que vale a pena ser resistida, pois a adaptação é quando muito uma pergunta, não uma resposta (p.243).” A definição baseada em contiguidade, por mais errada que fosse, ao menos tentava responder à pergunta de como podem surgir aproximações de comportamento ótimo ou adaptativo. Este artigo oferece uma resposta que evita a visão estreita dos conceitos de ordem, proximidade e fortalecimento e permite explicações plausíveis do comportamento tanto em laboratório como no dia-a-dia. Por ela omitir a ideia de fortalecimento, poder-se-ia duvidar que este mecanismo devesse ser chamado de “reforçamento”.
Alocação de Tempo entre Atividades
Vemos na Figura 1 que alimento periódico muda a alocação de tempo entre atividades, como a de bater asas e bicar ou beber e correr. Algumas atividades aumentam de frequência enquanto outras diminuem. Nenhuma noção de fortalecimento precisa entrar neste quadro. Pelo contrario, precisamos apenas perceber a mudança na alocação.
Estar vivo é se comportar, e podemos assumir que em uma hora de observação poderá ser observado uma hora válida de comportamento, assim como em um mês de observação poderá ser observado um mês válido de comportamento. Portanto, podemos comparar a alocação de comportamento a cortar uma pizza em pedaços. A Figura 2 ilustra essa ideia com um exemplo hipotético. O gráfico mostra a alocação do tempo no dia de um estudante típico. Aproximadamente 4 horas são gastas assistindo aula, 4 h estudando, 2,5 h se alimentando, 0,5 h no banheiro, 3 h em recreação e 2 h trabalhando. Isso tudo totaliza 16 h, sendo as outras 8 h usadas dormindo. Como um dia tem apenas 24 h, para que qualquer atividade aumente, outras têm que decrescer, e se alguma decrescer, outras têm que aumentar. Como veremos mais tarde, a finitude do tempo é importante na compreensão dos efeitos das contingências.
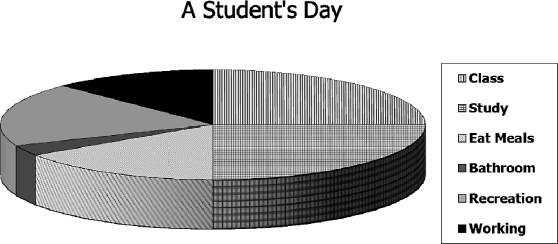
Fig. 2 Ilustração hipotética do conceito de alocação. As horas gastas em atividades por um estudante típico em um dia típico totalizam 16 h, as outras 8 h sendo usadas para dormir. Se mais tempo for gasto em uma atividade, menos será gasto em outras, e vice versa.
Medir o tempo gasto em uma atividade pode apresentar desafios. Por exemplo, quando exatamente está um pombo bicando um botão ou um rato pressionando uma barra? Ligar um dispositivo ao botão ou barra permite que se meça o tempo gasto com base no número de vezes em que o dispositivo é ativado. Por sorte, este método é em geral confiável; contar as vezes em que o dispositivo é ativado e por quanto tempo é em cada vez gera dados equivalentes (Baum, 1976). Apesar de no passado os pesquisadores muitas vezes acreditarem que contavam respostas discretas (bicadas e pressões), no presente ponto de vista estavam usando a ativação de dispositivos para medir o tempo gasto em atividades contínuas (bicar e pressionar).
Fora variações assistemáticas que podem ocorrer de amostra para amostra, a alocação do comportamento muda sistematicamente como resultado de duas fontes: indução e contingência. Como vemos na Figura 1, um evento ambiental indutor como alimento aumenta algumas atividades e, como o tempo é finito, necessariamente diminui outras. Por exemplo, alimento induz pombos a bicarem, choque elétrico induz agressão em ratos e macacos, e a remoção de alimento induz agressão em pombos. A Figura 3 mostra um exemplo hipotético. Acima (em Time 1) está representada a alocação de tempo entre atividades por um pombo em situação experimental que inclui um espelho e alimento periódico frequente. O pombo passa muito tempo se alimentando e descansando e pode direcionar algum comportamento agressivo ou reprodutivo para o espelho. Abaixo (Time 2) está representado o resultado do aumento do intervalo entre as entregas de alimento. Agora o pombo passa muito mais tempo apresentando comportamento agressivo para o espelho, menos tempo comendo, menos tempo descansando e pouco tempo em comportamento reprodutivo.
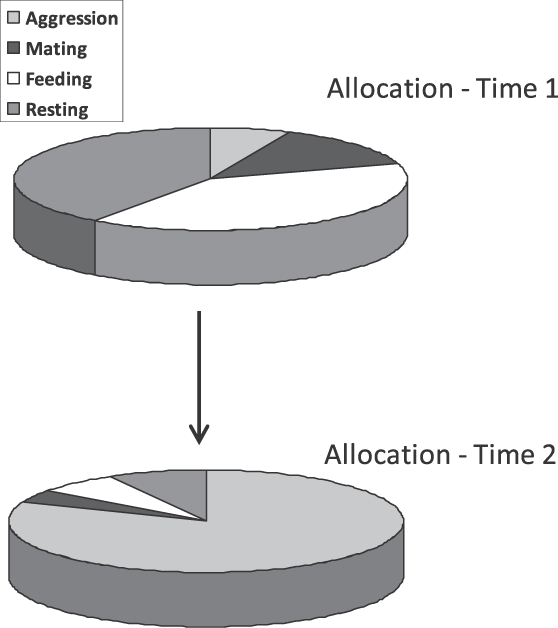
Fig. 3. Um exemplo hipotético ilustrando a mudança de comportamento devido a indução como mudança de alocação. Em Time 1, as atividades de um pombo em um espaço experimental apresenta Alocação 1 em reposta ao recebimento periódico de alimento. Em Time 2, o intervalo entre os recebimentos de alimento foi aumentado, resultando um aumento de agressão direcionado ao espelho. Outras atividades necessariamente diminuem.
Uma contingencia liga um evento ambiental a uma atividade, resultando em um aumento ou redução da atividade. Por exemplo, fazendo-se uma ligação entre comida e pressão à barra, aumenta-se o tempo gasto a pressionando; fazendo-se uma ligação entre choque elétrico e pressão a barra, geralmente diminui-se o tempo gasto pressionando. A Figura 4 ilustra com um exemplo hipotético. O diagrama de cima mostra a possível alocação de comportamento de uma criança em uma sala de aula. Muito tempo é gasto batendo em outras crianças e gritando, mas pouco tempo nas tarefas. Há tempo usado na interação com o professor, o que não tem relação com estar fazendo as tarefas. O diagrama de baixo mostra o resultado da atenção do professor ter se tornado contingente a se dedicar às tarefas. Agora a criança dedica muito tempo às tarefas, menos tempo batendo e gritando, e um pouquinho mais de tempo interagindo com o professor – provavelmente agora de uma forma mais amigável. Como o aumento de uma atividade diminui outra, uma contingencia positiva entre recompensa e uma atividade implica em uma contingencia negativa entre a recompensa e outras atividades – quanto mais gritos e batidas, menos atenção do professor. Uma forma de capturar ambos estes aspectos da contingência é considerar que a alocação inteira entra na contingência com recompensa (Baum, 2002).
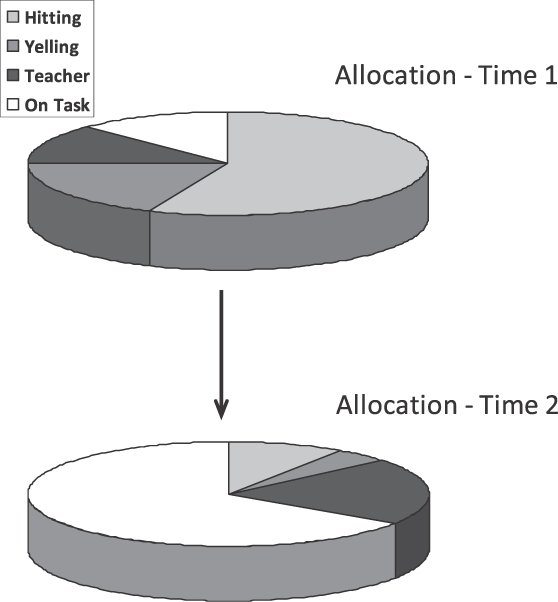
Fig. 4. Um exemplo hipotético ilustrando mudança de alocação por correlação. Em Time 1, as atividades de uma criança em sala de aula mostram Alocação 1, incluindo altas frequências de atividades inoportunas. Em Time 2, depois que a atenção do professor se tornou contingente a fazer as tarefas, Alocação 2 inclui mais tempo dedicado às tarefas e necessariamente menos tempo a atividades inoportunas.
Indução
O conceito de indução foi introduzido por Evalyn Segal em 1972. Para definir, ela se baseou nas definições de dicionário “estimular a ocorrência de” e “ocasionar,” e comentou “ela implica em uma certa indireção em causar algo a acontecer, e parece apta para falar sobre operações que podem ser eficazes apenas em conjunção com outros fatores” (p.2). Na visão molar, essa “indireção” é crucial, pois o conceito de indução significa que, em um dado contexto, a mera ocorrência de certos eventos, como alimento – i.e. indutores – resulta na ocorrência ou aumento de tempo dedicado a certas atividades. Todas as várias atividades que em algum ponto foram chamadas de “interinas,” “terminais,” “facultativas,” ou “adjuntivas,” como as apresentadas na Figura 1, podem ser agrupadas como atividades induzidas (Hinline, 1984). São induzidas em qualquer situação que inclui eventos como alimento ou choque elétrico, os mesmos eventos que previamente foram chamados de “reforçadores,” “punidores,” “estimulos incondicionais”, ou “libertadores.” A indução é distinta da ideia mais estreita de eliciação, que presume uma relação temporal próxima entre estimulo e resposta. Um motivo para a introdução de um termo novo, mais amplo é que os indutores não precisam de uma relação temporal próxima às atividades as quais induzem (Figura 1). Sua ocorrência em uma dado ambiente é o suficiente para induzir atividades, a maioria das quais estão estreitamente relacionadas ao evento indutor nas interações da espécie com seu ambiente – alimento induz a atividades relacionadas a alimento (por exemplo pombos a bicarem), choque elétrico induz a atividades ligadas a dor (como agressividade e corrida), e uma possível companheira induz a cortejo e cópula.
Indução e Controle de Estímulo
A indução se assemelha ao conceito de controle de estímulos de Skinner. Um estímulo discriminativo não se constitui de uma relação temporal um-para-um com o responder, mas sim aumenta a taxa de tempo gasta na atividade em sua presença. Em termos presentes, um estímulo discriminativo modula a alocação de atividades em sua presença; ele induz uma certa alocação de atividades. Pode-se dizer que um estímulo discriminativo estabelece a ocasião ou contexto para uma atividade operante. De forma similar, pode-se dizer que um indutor pode ocasionar ou criar um contexto para a atividade induzida. O controle de estímulos pode às vezes parecer mais complicado, como em uma discriminação condicional que requer uma certa combinação de eventos, mas não importa o quão complexa seja a discriminação, a relação com o comportamento é a mesma. De fato, embora nós concebamos o controle de estímulo como um produto da história de vida do indivíduo (ontogenia), se nós aceitarmos a ideia de que algumas instâncias do controle de estímulos podem existir como resultado da filogenia, então o controle de estímulos e a indução seriam dois termos diferentes para o mesmo fenômeno: o efeito do contexto na alocação comportamental. A seguir, assumirei que a indução ou o controle de estímulos podem surgir tanto como resultados da filogenia ou da história de vida.
Apesar de Pavlov ter pensado em termos de reflexos e medido apenas secreção estomacal ou salivação, hoje podemos aplicar os termos atuais retrospectivamente e dizer que ele estava estudando indução (Hineline, 1984). Zener (1937), repetindo o procedimento de Pavlov com cães não restritos, observou que durante a execução de um som que precedia a comida, os cães se aproximariam da vasilha de comida, balançariam suas caudas, e assim por diante – tudo isso sendo comportamento induzido. Os experimentos de Pavlov nos quais estímulos condicionais como sons e imagens eram correlacionados com indutores como comida, choque elétrico e ácido na boca, mostraram que correlacionando um estímulo neutro com um indutor muda-se a função do estímulo. Nos termos atuais, ele se torna um estímulo indutor condicional ou um indutor condicional, equivalente a um estímulo discriminativo. De forma semelhante, atividades ligadas por contingência a indutores se tornam atividades relacionadas a indutores. Por exemplo, Shapiro (1961) descobriu que quando a pressão à barra por um cão produzia comida, o cão salivava sempre que esta era pressionada. Nós desenvolveremos essa ideia subseqüentemente, quando discutirmos os efeitos das contingências.
Eventos Filogeneticamente Importantes
Por adquirirem a capacidade de induzir como o resultado de muitas gerações de seleção natural – da filogenia – eu os chamo de Eventos Filogeneticamente Importantes (PIEs; Baum, 2005). Um PIE é um evento que afeta diretamente a sobrevivência e reprodução. Alguns PIEs aumentam as chances de sucesso reprodutivo por sua presença; outros reduzem-no. Entre os eventos que aumentam, temos comida, abrigo e oportunidades de acasalamento; entre os eventos que reduzem, temos exemplos como predadores, parasitas e condições climáticas severas. Os indivíduos para os quais esses eventos não eram importantes produziram menos descendentes sobreviventes que seus competidores – para os quais eles eram importantes – e não estão mais representados na população.
Os PIEs são muitos e variados, dependendo do ambiente em que uma espécie evoluiu. Nos humanos, eles incluem eventos sociais como sorrisos, expressões zangadas e contato visual, eventos que afetam a aptidão por causa de nossa longa história de vivência em grupos e pela importância do pertencimento a um grupo para sobrevivência e reprodução. PIEs incluem também eventos perigosos e amedrontadores como ferimentos, lugares altos, cobras, e objetos grandes se aproximando rapidamente.
De acordo com sua origem evolutiva, PIEs e seus efeitos dependem da espécie. Breland e Breland (1961) relataram a intrusão de atividades induzidas em suas tentativas de treinar diversas espécies usando contingências com comida. Porcos procuravam por moedas de madeira da mesma forma que procurariam por comida; guaxinins limpavam moedas como se eles estivessem limpando comida; galinhas ciscavam o chão no mesmo espaço que elas eram alimentadas. Quando a relevância da filogenia foi amplamente reconhecida, artigos e livros começaram a apontar para “restrições biológicas” (Hinde & Stevenson-Hinde, 1973; Seligman, 1970). Em humanos, para as reações a eventos como comida e ferimentos, nós podemos acrescentar a indução de respostas especiais para indivíduos da mesma espécie conhecidos ou desconhecidos, como o sorriso, o levantamento de sobrancelha, o mostrar de língua (Eibl-Eibesfeldt, 1975).
Relatos de reforçadores e punidores que não fazem referência à história evolutiva falham em explicar por que esses eventos são efetivos da forma que são. Por exemplo, Premack (1963; 1965) propôs que o reforçamento pode ser pensado como uma contingência na qual a disponibilidade de uma atividade de “alta probabilidade” depende da ocorrência de uma atividade de “baixa probabilidade”. Timberlake e Allison (1974) elaboraram essa ideia acrescentando que qualquer atividade restrita a ocorrer abaixo de seu nível de linha de base, quando está livremente disponível, está em “privação de resposta” e, quando feita contingente a qualquer atividade com um nível mais baixo de privação, reforçará essa atividade. Seja qual for a validade dessas generalizações, elas requerem o questionamento “Por que os níveis de linha de base de atividades como comer e beber são mais altos que os níveis de linha de base de correr e pressionar a barra?” Privação e fisiologia não são respostas, a questão permanece, “Porquê os organismos são constituídos de tal forma que quando a comida (ou água, abrigo, sexo, etc. ) está em estado de privação, ele se torna um potente indutor?” Ao ligar a importância maior de comer e beber (comida e água) à filogenia explica-se porquê esses eventos são importantes em um sentido último, e que o conceito de indução explica como eles são importantes para a flexibilidade do comportamento.
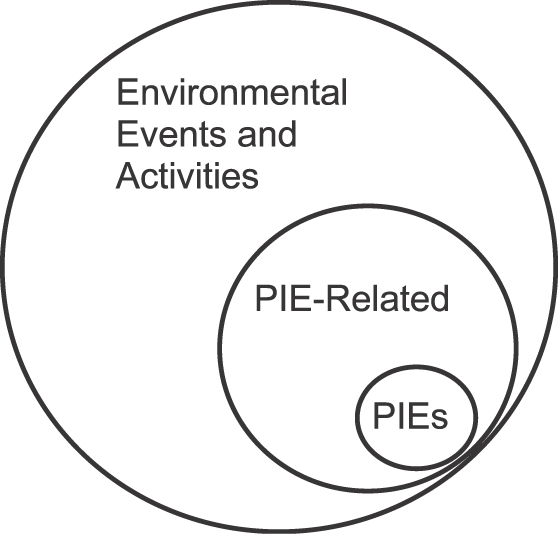
Fig. 5. Diagrama ilustrando que Eventos Filogeneticamente Importantes (PIEs) constituem um subconjunto de eventos relacionados aos PIEs, que por sua vez são um subconjunto de todos os eventos.
A Figura 5 ilustra que, dentre todos os eventos possíveis – estímulos ambientais e atividades – simbolizados pelo círculo maior, PIEs são um pequeno subconjunto, e estímulos e atividades relacionados aos PIEs são um subconjunto maior. O fato de o subconjunto de estímulos e atividades relacionadas aos PIEs incluir o subconjunto de PIEs indica que os próprios PIEs geralmente são do tipo relacionados a PIEs, porquê ambientes naturais e situações de laboratório mostram que a ocorrência de um PIE como comida prediz como e quando mais comida estará disponível. Comida geralmente prediz mais comida, e a presença de um predador geralmente prediz perigo contínuo.
Um conceito chave sobre induções é que um PIE induz qualquer atividade relacionada ao PIE. Se um PIE, como comida, é feito contingente a uma atividade como pressão à barra, a atividade (pressão à barra) se torna relacionada a um PIE (relacionado à comida). A base experimental para esse conceito pode ser encontrada em experimentos que mostram que PIEs funcionam como estímulos discriminativos. Existem muitos experimentos deste tipo, e eu ilustrarei com três exemplos mostrando que comida contingente a uma atividade induz essa atividade.
Bullock e Smith (1953) treinaram ratos por 10 sessões. Cada sessão consistiu em 40 pelotas de comida, cada uma produzida por uma pressão à barra (isto é, Razão Fixa 1) seguidas por 1 hora de extinção (pressão à barra ineficaz). A figura 6 mostra o número médio de pressões durante a hora de extinção durante as dez sessões. O decréscimo se assemelha ao decréscimo em “erros” que ocorrem durante a formação de uma discriminação. Foi assim que Bullock e Smith interpretaram seus resultados: O estímulo positivo (SD) foi pressão-mais-comida, e o estímulo negativo (SΔ) foi pressão-sem-comida. A comida foi o elemento crítico controlando o responder. Sendo assim, os resultados apoiam a ideia de que a comida funciona como um estímulo discriminativo.

Fig. 6. Resultados de um experimento no qual a própria comida serviu como estímulo discriminativo. O número de respostas durante uma hora de extinção seguindo 40 respostas que produziram comida decresceu durante 10 sessões de experimento. As respostas seriam análogas a “erros”. Os dados foram publicados em um artigo por Bullock e Smith (1953).
O segundo achado ilustrativo, relatado por Reid (1958), é referido como “restabelecimento” e tem sido estudado extensivamente (por exemplo, Ostlund & Balleine, 2007). A figura 7 mostra o resultado de forma esquemática. Reid estudou ratos, pombos e estudantes. Primeiro, ele treinou o responder ao seguir cada resposta com comida (ratos e pombos) ou uma ficha (estudantes). Uma vez que o responder foi bem estabelecido, ele o submeteu no primeiro dia a 30 minutos de extinção. Isso foi repetido nos dias 2 e 3. Então, no dia 3, quando o responder desapareceu, ele apresentou um pouco de comida (ratos e pombos) ou uma ficha (estudantes). Essa recompensa gratuita foi imediatamente seguida por um aumento rápido no responder (circulado na figura 7). Assim, a comida ou ficha dada gratuitamente reinstalou o responder, funcionando da mesma forma que um estímulo discriminativo.
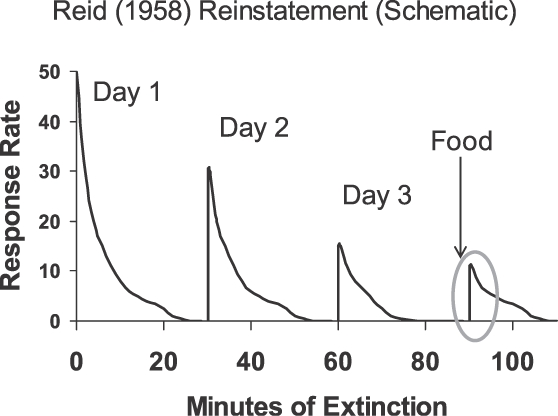
Fig. 7. Imagem dos resultados de Reid (1958) com o restabelecimento do responder seguindo a extinção depois de uma única apresentação do evento originalmente contingente (comida para ratos e pombos; fichas para estudantes) de forma independe da resposta. Um aumento rápido no responder (circulado) seguiu a apresentação, sugerindo que o evento contingente funcionou como um estímulo discriminativo ou um indutor.
Como uma terceira ilustração, nós temos dados que o próprio Skinner apresentou em seu artigo de 1948 sobre “superstição”. A figura 8 reproduz o registro cumulativo que Skinner diz ter mostrado o “recondicionamento” de uma resposta de pular de um lado ao outro por reforço adventício. Olhando de perto para as duas primeiras ocorrências de comida (setas circuladas), entretanto, nós vemos que nenhuma resposta ocorreu imediatamente antes da primeira ocorrência e nem possivelmente imediatamente antes da segunda ocorrência. O pular seguiu a comida; ele não o precedeu. Os resultados de Skinner mostram restabelecimento, não reforçamento. Em outras palavras, o registro mostra a indução por comida de forma periódica similar àquela mostrada na figura 1.
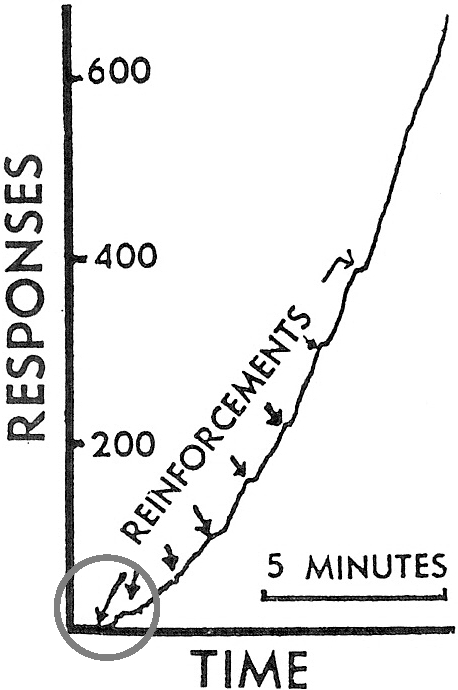
Fig. 8. Registro cumulativo que Skinner (1948) apresentou como evidência do “recondicionamento” de uma resposta de pular de um lado para o outro. A comida foi proporcionada a cada minuto independente do comportamento do pombo. As disponibilizações de comida (rotuladas “reforçadores”) são indicadas por setas. Nenhuma resposta precedeu imediatamente a primeira e possivelmente a segunda disponibilização de comida (circuladas), indicando que o resultado é, de fato, um exemplo de restabelecimento. Copyright 1948 pela American Psychological Association. Reproduzida por permissão.
Todos os três exemplos mostram que a comida induz atividades “operantes” ou “instrumentais” que estão correlacionadas com ela. O PIE induz atividades relacionadas a ele, e esses exemplos mostram que atividades relacionadas ao PIE incluem atividades operantes que estiverem correlacionadas com ele através de contingência. Contrária à visão corrente de reforçadores – que sua função primária é fortalecer a resposta à qual eles são contingentes, mas que eles têm um papel secundário como estímulos discriminativos – a visão presente afirma que existe apenas a função do estímulo. Nenhuma noção de “fortalecimento” ou “reforçamento” precisa ser levada em conta.
Uma forma de entender o poder indutor de um PIE como a comida é reconhecer que na natureza e no laboratório a comida geralmente é um sinal de que mais comida estará presente naquele ambiente para aquela atividade. Um pombo ciscando por sementes de grama, ao achar uma semente, tem uma probabilidade maior de achar mais sementes se continuar ciscando aquele local (“busca focal”; Timberlake & Lucas, 1989). Uma pessoa que está buscando um encontro na internet, ao encontrar uma perspectiva esperançosa em um site, é provável de encontrar mais se continuar buscando naquele site. Por essa razão, Michael Davidson e eu sugerimos que a metáfora do fortalecimento deveria ser substituída pela metáfora de orientação (Davidson & Baum, 2006).
Um artigo por Gardner e Gardner (1988) argumentou vigorosamente em favor de um papel maior ao processo de indução em explicar as origens do comportamento, sugerindo até mesmo que a indução é sua principal determinação. Eles chegaram a essa conclusão a partir de estudos de filhotes de chimpanzés criados por humanos. A indução pareceu explicar muito do comportamento dos chimpanzés. Mas apenas a indução não pode explicar a seleção entre atividades, porque devemos também explicar a ocorrência dos PIEs que induzem a várias atividades. Isso requer que consideremos o papel da contingência, porque as contingências governam a ocorrência de PIEs. O relato dos Gardner não dá nenhuma explicação para a flexibilidade do comportamento dos indivíduos, porque eles negligenciaram a importância da contingência dentro da evolução comportamental (Baum, 1988).
Analogia com a Seleção Natural
Como mencionado anteriormente, a otimização por si só não é uma explicação, mas requer explicação por um mecanismo. A evolução de uma população por seleção natural oferece um exemplo. A evolução darwiniana requer três ingredientes: (1) variação; (2) recorrência; e (3) seleção (por exemplo, Dawkins, 1989; Mayr, 1970). Quando estão todos presentes, a evolução acontece; eles são necessários e suficientes. Os fenótipos em uma população variam em seu sucesso reprodutivo – ou seja, no número de descendentes sobreviventes que eles produzem. Os fenótipos tendem a recorrer de geração em geração – isto é, descendentes tendem a se parecer com seus pais – porquê os pais transmitem material genético para seus descendentes. A seleção ocorre devido à capacidade do ambiente de suportar uma população impor um limite para seu tamanho, e como conseqüência muitos descentes não sobrevivem, e o fenótipo com mais descendentes sobreviventes aumenta naquela população.
A evolução do comportamento em um curso de vida (ou seja, modelagem), visto como um processo darwiniano, também requer variação, recorrência e seleção (Staddon & Simmelhag, 1971). A variação ocorre nas atividades em si e entre atividades. Já que cada atividade é composta de partes que são também atividades, a variação em uma atividade é também variação entre atividades, mas em uma escala menor de tempo (Baum, 2002; 2004). PIEs e estímulos relacionados a PIEs induzem várias atividades em qualquer situação. Portanto, a indução é o mecanismo de recorrência, fazendo as atividades persistirem no tempo, de forma análoga à reprodução que permite que os fenótipos persistam em uma população com o passar do tempo. A seleção ocorre por causa da finitude do tempo, o que é análogo ao limite do tamanho da população definido pela capacidade do ambiente de suportar essa população. Quando os PIEs induzem várias atividades, essas atividades competem entre si, uma vez que o aumento em uma exige que outras diminuam. Por exemplo, quando o pombo é treinado para bicar teclas que ocasionalmente produzem comida, a bicada é induzida por comida, mas o bicar compete pelo tempo com outras atividades de segundo plano como ficar se arrumando e descansar (Baum, 2002). Se a tecla da esquerda produz comida cinco vezes mais que a tecla da direita, então a combinação de tecla da esquerda e comida induz mais bicadas na tecla da esquerda do que bicadas na tecla da direita – a base para a relação de pareamento (Herrnestein, 1961; Davidson & McCarthy, 1988). Devido ao limite superior no bicar determinado por sua competição com atividades de segundo plano, o bicar na tecla da esquerda compete por tempo com o bicar na tecla da direita (Baum, 2002). Se nós calcularmos a taxa de bicadas na tecla da esquerda pela taxa de bicadas na tecla da direita e as compararmos com a taxa de comida conseguida com esquerda pela taxa conseguida pela direita, o resultado pode variar do pareamento estrito entre taxa de bicada e taxa de comida entre as duas teclas (Baum, 1974; 1979), mas os resultados geralmente se aproximam do pareamento, e simples evidências matemáticas mostram o pareamento como sendo ideal (Baum, 1981b).
Sendo assim, embora a evolução de populações de organismos e a evolução comportamental possam ser vistas como processos darwinianos otimizantes, elas funcionam por mecanismo próprios. O resultado é a aproximação da otimização, mas os mecanismos explicam esse resultado.
Para compreender a evolução comportamental completamente, nós precisamos não apenas entender os efeitos de competição dos PIEs, mas nós precisamos levar em consideração a ocorrência dos PIEs – como eles são produzidos pelo comportamento. Nós precisamos incluir os efeitos das contingências.
Contingências
Sempre se soube que a noção de reforço é incompleta. Por si mesma, ela não inclui nenhuma explicação da origem do comportamento a ser fortalecido; o comportamento precisa ocorrer antes que possa ser reforçado. Segal (1972) pensou que talvez a indução pudesse explicar a origem do comportamento operante. Talvez a contingência selecione comportamento originalmente induzido. Antes de seguir essa ideia, precisamos estar esclarecidos sobre o significado do termo “contingência”.
Nas citações que vimos anteriormente, Skinner definiu contingência como contigüidade – um reforçador precisaria apenas seguir a uma resposta. Contrária a essa visão baseada em “ordem e proximidade”, uma contingência não é uma relação temporal. Rescorla (1968; 1988) pode ter sido o primeiro a apontar que apenas a contigüidade não pode especificar uma contingência, porquê uma contingência requer uma comparação entre pelo menos duas ocasiões diferentes. A figura 9 ilustra o fato com uma tabela 2 por 2, na qual as colunas mostram a presença e ausência do Evento 1, e e as linhas mostram a presença e ausência do Evento 2. O evento 1 poderia ser um som ou bicada na tecla; o evento 2 poderia ser comida ou choque elétrico. A célula superior esquerda representa a conjunção entre esses dois eventos (isto é, contigüidade); a célula inferior direita representa a conjunção da ausência dos dois eventos. Essas suas conjunções (marcas) devem ambas ter alta probabilidade para uma contingência ocorrer. Se, por exemplo, a probabilidade do Evento 2 fosse a mesma, independente da presença ou ausência do Evento 1 (células superior esquerda e superior direita), então nenhuma contingência existiria. Portanto, a presença de uma contingência requer a comparação entre duas ocasiões temporalmente separadas (Evento 1 presente e evento 1 ausente). Uma correlação entre a taxa de uma atividade e a taxa de comida implicaria em mais de duas comparações – por exemplo, notando várias taxas de comida durante várias taxas de bicadas (a base de uma função de feedback; Baum, 1973, 1989). Contrário a ideia de que a contingência requer apenas conjunção temporal, a figura 9 mostra que contingências acidentais deveriam ser raras, porquê uma contingência acidental iria requerer pelo menos duas conjunções acidentais.
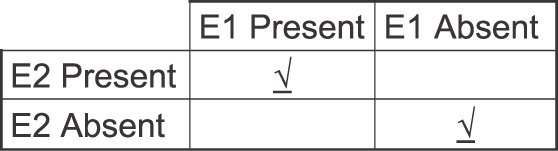
Fig. 9. Porquê uma contingência ou correlação não é simplesmente uma relação temporal. A tabela 2 por 2 mostra que as conjunções possíveis da presença e ausência de dois eventos, E1 e E2. Uma contingência positiva existe entre E1 e E2 somente se duas conjunções ocorrem com alta probabilidade entre em tempos diferentes: a presença de ambos os eventos e ausência de ambos os eventos (indicadas por marcas). A conjunção dos dois eventos isoladamente (contigüidade) não é suficiente.
Não é que as relações temporais sejam completamente irrelevantes, mas apenas que elas são relevantes de forma diferente daquela que uma concepção do reforço tradicional iria requerer. Considerando que as formulações de Skinner atribuíram um papel direto à contiguidade, a concepção de contingência ilustrada na figura 9 sugere em vez disso que o efeito da contiguidade é indireto. Por exemplo, inserir atrasos não sinalizados em uma contingência geralmente reduz a taxa de atividade (por exemplo, bicada na tecla) que produz o PIE (por exemplo, comida). Esses intervalos devem afetar o grau da correlação. Uma medida de correlação como o coeficiente de correlação (r) deveria decrescer ao passo que o atraso médio aumentasse. Atrasos afetam a clareza de uma contingência como aquela da figura 9, e afetam a variância em uma correlação (para futuras discussões, ver Baum, 1973).
Esquiva
Nenhum fenômeno ilustra melhor a inadequação da visão molecular do comportamento do que a esquiva, porque toda a função da esquiva é que quando a resposta ocorre, nada se segue. Por exemplo, Sidman (1953) treinou ratos em um procedimento no qual a pressão à barra adiava choques elétricos que, na ausência de pressão, ocorriam a uma taxa regular. Quanto maior a taxa de pressão à barra, menor a taxa de choque, e se a pressão ocorresse a uma taxa suficientemente elevada, a taxa de choque era reduzida a um valor próximo de zero. Para tentar explicar a pressão à barra, teorias moleculares recorrem ao “medo” invisível eliciado por estímulos invisíveis, a redução do “medo” resultante de cada pressão à barra – então chamada de teoria dos “dois fatores” (Anger, 1963; Dinsmoor, 2001). Em contraste, uma teoria molar de esquiva não faz referência a variáveis ocultas, mas se baseia na redução mensurável na freqüência de choque resultante da pressão à barra (Herrnstein, 1969; Sidman, 1953; 1966). Herrnstein e Hineline (1966) testaram a teoria molar diretamente através do treinamento de ratos em um procedimento onde a pressão à barra reduzia a freqüência do choque, mas não podia reduzi-la à zero. Uma explicação molecular requer postular uma redução invisível do medo que depende unicamente da redução da taxa de choque, admitindo implicitamente a explicação molar (Herrnstein, 1969).
Em nossos termos, a contingência negativa entre a taxa de choque e taxa de pressão à barra resulta em tornar a resposta de pressão à barra uma atividade relacionada ao choque. Choques, então, induzem a pressão à barra. No procedimento de Herrnstein-Hineline, esta indução é clara, porque o choque nunca desaparece por completo, mas no procedimento de Sidman, podemos supor que o contexto experimental – câmara, barra, etc – também induz a pressão à barra, como estímulos discriminativos ou indutores condicionais. Muitas evidências apóiam essas idéias. Choques extras não contingentes a resposta aumentam a freqüência de respostas de pressão à barra (Sidman, 1966; cf. restabelecimento discutido anteriormente); a extinção da esquiva de Sidman é prolongada quando a taxa de choque é reduzida a zero, mas torna-se mais rápida na repetição (Sidman, 1966; cf. o estudo de Bullock & Smith discutido anteriormente); a extinção na esquiva de Herrnstein-Hineline na qual a pressão à barra passa a não reduzir a taxa de choques depende da quantidade de redução da taxa de choque durante o treinamento (Herrnstein & Hineline, 1966); as sessões de esquiva de Sidman geralmente começam com um período de “aquecimento”, no qual choques são apresentados até que inicie a pressão à barra – isto é, até que o choque induza a pressão à barra (Hineline, 1977).
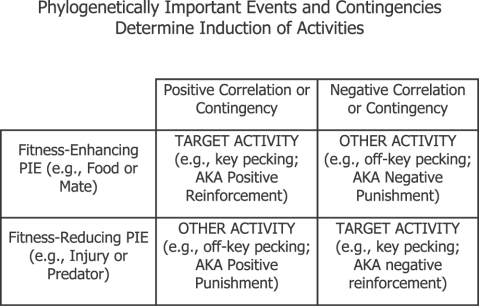
Fig. 10. Diferentes correlações ou contingências induzem ou a atividade alvo ou atividades outras que não a alvo, dependendo se o Evento Filogeneticamente Importante (PIE) envolvido normalmente aumenta ou diminui a aptidão (sucesso reprodutivo) por sua presença.
A Figura 10 ilustra por que o choque se torna um estímulo discriminativo ou um indutor condicional nos procedimentos de esquiva. Esta tabela dois-por-dois mostra os efeitos de contingências positivas e negativas (colunas) pareados com PIEs que elevam ou diminuem a aptidão (linhas). A seleção natural garante que PIEs perigosos (redutores de aptidão, como lesões, doenças, ou predadores) induzam atividades defensivas (por exemplo, esconder-se, paralisar-se, ou fugir) que removem ou atenuam o perigo. (Aqueles indivíduos da população que não conseguiram se comportar confiavelmente de tal modo geraram menos descendentes sobreviventes.) Na esquiva, uma vez que choque ou lesão são redutores de aptidão, qualquer atividade que possa evitá-los será induzida por eles. Após o treino de esquiva, a atividade operante (por exemplo, pressão à barra) torna-se uma atividade de manutenção de aptidão defesiva (condicional), e é induzida juntamente com outras atividades relacionadas ao choque; o choque em si e a (perigosa) câmara operante fazem a indução. A célula inferior direita na tabela corresponde às relações tipicamente chamadas de “reforçamento negativo”.
A seleção natural garante também que os PIEs que elevam a aptidão (como, presas, abrigo, ou um companheiro) induzam atividades de aumento de aptidão (por exemplo, alimentar-se, abrigar-se, ou cortejar). (Indivíduos na população que se comportaram confiavelmente de tal modo deixaram mais descendentes sobreviventes.) Na célula superior esquerda na Figura 10, um PIE que eleve a aptidão (por exemplo, alimento) está em uma relação positiva com uma atividade alvo (operante), e o treinamento resulta na indução da atividade alvo juntamente com outras atividades de melhoria de aptidão relacionadas a PIEs. Esta célula corresponde a relações tipicamente chamadas de “reforçamento positivo”.
Quando a resposta de pressão à barra é relacionada com comida ou quando é relacionada com choque, a comida ou o choque funcionam como estímulo discriminativo, induzindo a pressão à barra. A comida prediz mais comida; o choque prediz mais choques. Uma correlação positiva entre a taxa de comida e a taxa de pressão à barra gera a condição para que a pressão à barra se torne uma atividade relacionada à comida. A correlação negativa entre a taxa de choque e taxa de pressão à barra gera a condição para que a pressão à barra se torne uma atividade relacionada com o choque. Mais precisamente, as correlações criam as condições para a comida e o choque induzirem alocações que incluam quantidades substanciais de tempo gasto pressionando à barra.
Nas outras duas células da figura 10, a atividade alvo iria ou produzir um PIE de redução de aptidão – a célula tipicamente chamada de “punição positiva” – ou evitar um PIE de aumento de aptidão – célula normalmente chamada de “punição negativa”. De qualquer maneira, a atividade alvo reduziria a aptidão e seria impedida de fazer parte de outras atividades relacionadas à PIEs. Em vez disso, outras atividades, incompatíveis com a atividade alvo, que manteriam a aptidão, são induzidas. Experimentos com punição positiva configuram um conflito, porque a atividade alvo geralmente produz tanto PIE de aumento de aptidão quanto PIE de diminuição de aptidão. Por exemplo, quando a atividade operante (como, pombos bicando uma tecla) produz tanto comida como choque elétrico, é comum que a atividade se reduza a níveis abaixo do seu nível na ausência de choque (Azrin & Holz, 1966; Rachlin & Herrnstein, 1969). A comida induz a resposta de bicar, mas outras atividades (por exemplo, bicar fora da tecla) são negativamente correlacionadas com a taxa de choque e são induzidas pelo choque. O resultado é uma alocação de compromisso incluindo menos bicadas na tecla. Chegaremos a uma conclusão semelhante sobre punição negativa, em que a atividade alvo (por exemplo, bicar) cancela um PIE de aumento de aptidão (apresentação de comida), quando discutirmos auto-preservação negativa. A ideia de que a punição induz atividades alternativas à atividade punida é ainda apoiada pela observação de que se a situação inclui outra atividade que é positivamente correlacionada com a comida e não produz nenhum choque, essa atividade predominará (Azrin & Holz, 1966).
Os efeitos da Contingência
A contingência relaciona uma atividade a um evento indutor e altera a alocação de tempo entre as atividades pelo aumento do tempo gasto nas atividades relacionadas. O aumento no tempo gasto em uma atividade – digamos, pressão à barra – quando a comida é feita contingente a ela foi no passado atribuído ao reforço, mas os nossos exemplos sugerem que seja devido à indução. O aumento nas respostas de pressão à barra resulta da combinação de contingência e indução, porque a contingência transforma a pressão à barra em uma atividade relacionada à comida (relacionada a PIE), como mostrado nos experimentos de Bullock e Smith (1953) (Figura 6) e de Reid (1958) (Figura 7). Podemos resumir essas idéias da seguinte forma:
- Eventos filogeneticamente importantes (PIEs) são indutores incondicionais.
- Um estímulo correlacionado com um PIE torna-se um indutor condicional.
- Uma atividade correlacionada positivamente com um PIE que aumente a aptidão torna-se uma atividade relacionada a PIE condicional induzida – normalmente chamada de “operante” ou atividade “instrumental”.
- Uma atividade negativamente correlacionada com um PIE redutor de aptidão torna-se uma atividade relacionada a PIE condicional induzida – normalmente chamada de “esquiva operante”.
- Um PIE induz atividade operante relacionada a ele.
- Um indutor condicional induz atividade operante relacionada ao PIE ao qual está relacionado.
Os efeitos da contingência não precisam incluir nenhuma noção de fortalecimento ou reforço. Considere o exemplo da resposta de pressão à barra mantida por choque elétrico. A Figura 11 mostra um exemplo de registro cumulativo da pressão à barra de um macaco-de-cheiro do estudo de Malagodi, Gardner, Ward, and Magyar (1981). As curvas assemelham-se às produzidas por um esquema de intervalo fixo de comida, começando com uma pausa e, em seguida, acelerando a uma taxa de resposta maior até ao fim do intervalo. No entanto, neste registro a última pressão à barra do intervalo produz um choque elétrico. O resultado requer, contudo, que o macaco seja previamente treinado para pressionar à barra para evitar o choque elétrico. Embora no treino de esquiva o choque funcione como um estímulo aversivo ou punidor, na Figura 11 atua, paradoxalmente, para manter a resposta de pressão à barra, como se fosse um reforçador. O paradoxo é resolvido quando reconhecemos que o choque, como um PIE, induz a pressão à barra porque o treino de esquiva feito anteriormente tornou a resposta de pressão à barra uma atividade relacionada ao choque. Nenhuma noção de reforço atua – apenas o efeito da contingência. O choque induz a pressão à barra, a pressão à barra produz o choque, o choque induz a pressão à barra, e assim por diante, em um ciclo.
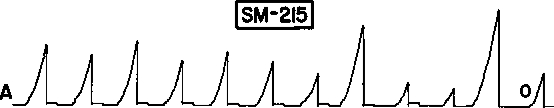
Fig. 11. Registro cumulativo de um macaco-de-cheiro pressionando uma barra que produz um choque elétrico, no final do intervalo fixo. Mesmo que o macaco tenha sido previamente treinado para evitar o mesmo choque, ele agora continua a pressionar quando o pressionar produz o choque. Este resultado aparentemente paradoxal é explicado pela visão molar do comportamento como um exemplo de indução. Reproduzido de Malagodi, Gardner, Ward, e Magyar (1981).
Os diagramas da Figura 12 mostram como a contingência fecha um ciclo. O diagrama à esquerda ilustra somente a indução. O ambiente E produz um estímulo S (choque), e o organismo O produz as atividades induzidas B. O diagrama à direita ilustra o efeito de contingência. Agora, o ambiente E liga o comportamento B ao estímulo S, resultando em um ciclo de indução e contingência.
O leitor já deve ter percebido que a mesma situação e o mesmo diagrama se aplicam às experiências com outros PIEs, como comida. Por exemplo, a Figura 1 mostra que, entre outras atividades, o alimento induz o bicar em pombos. Quando uma contingência funciona de forma que bicadas também produzam alimentos, temos o mesmo tipo de ciclo como mostrado na Figura 12, exceto que o estímulo S é a comida e o comportamento B é bicar. A comida induz a atividade, e a atividade (operante) produz o alimento, o que induz a atividade, e assim por diante, como mostrado pelos resultados de Bullock e Smith (1953) (Figura 6) e do efeito de restabelecimento de Reid (1958) (Figura 7).
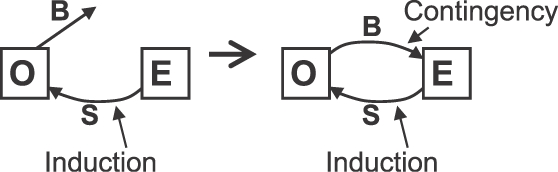
Fig. 12. Maneira pela qual a contingência completa um ciclo no qual uma atividade operante (B) produz um evento indutor (S), que por sua vez induz mais da atividade. “O” representa organismo. “E” representa o ambiente. Esquerda: somente indução. Direita: a contingência fecha o ciclo. Indução ocorre porque a atividade operante é ou se torna relacionada com o indutor (S; um PIE).
A Figura 13 mostra os resultados de um experimento que ilustra os efeitos de contingências positivas e negativas (Baum, 1981a). Pombos treinados a bicar uma tecla de resposta que, ocasionalmente, produz comida, foram expostos a sessões diárias em que o cronograma de recompensa começava com uma correlação positiva entre a taxa de bicar e a de comida. Era um tipo de esquema de intervalo variável, porém era necessária apenas uma ocorrência do bicar em qualquer momento do intervalo programado para que a comida fosse apresentada ao final desse intervalo (tecnicamente, um esquema conjunto de intervalo variável de 10 s e razão fixa 1). Num ponto imprevisível na sessão, a correlação mudava para negativa; agora, o bicar durante um intervalo programado cancelava a apresentação de comida ao final do intervalo. Finalmente, a um ponto imprevisível, a correlação revertia para positiva. A Figura 13 mostra quatro registros acumulados representativos de 4 pombos deste experimento. Inicialmente, o bicar ocorria a uma taxa moderada, em seguida, diminuiu quando a correlação mudou para negativo (primeira linha vertical), e depois aumentou novamente quando a correlação reverteu-se para positivo (segunda linha vertical). A efetividade da contingência negativa variou entre os pombos; suprimiu completamente o bicar a tecla no Pombo 57, e relativamente pouco no Pombo 61. Seguindo a nossa linha presente, estes resultados sugerem que quando a correlação era positiva, a comida induzia o bicar a tecla a uma taxa mais elevada do que quando a correlação foi negativa. De acordo com os resultados mostrados na Figura 1, no entanto, espera-se que a comida continue induzindo as bicadas. Como esperamos a partir da célula superior direita da Figura 10, face a uma contingência negativa, pombos tipicamente bicam fora da tecla, por vezes, bem próximo a ela. A correlação negativa entre a comida e o bicar a tecla constitui uma correlação positiva entre a comida e o bicar fora da tecla; a atribuição entre as duas atividades se altera nas três fases mostradas na Figura 13.
Os resultados na fase do meio dos registros na Figura 13 se assemelham a auto-manutenção negativa (Williams & Williams, 1969). Em auto-modelagem, um pombo é repetidamente apresentado a uma breve luz em uma tecla, que é seguida por comida. Cedo ou tarde, a luz passa a induzir bicadas na tecla, e o bicar a tecla iluminada se torna persistente; auto-modelagem torna-se auto-manutenção. Na auto-manutenção negativa, bicar a tecla cancela a apresentação de comida. Esta contingência negativa provoca uma redução no bicar a tecla, como na Figura 13, ainda que não o suprima totalmente (Sanabria, Sitomer & Killeen, 2006). Presumivelmente, um comprometimento ocorre entre o bicar na tecla e o bicar fora da tecla – a atribuição de tempo entre o bicar na tecla e bicar fora da tecla muda de um sentido para o outro. Possivelmente, a luz da tecla pareada a comida induz o bicar na tecla, enquanto que o alimento induz o bicar fora da tecla.
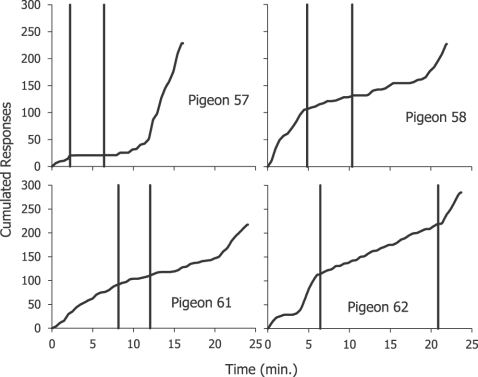
Fig. 13. Resultados de um experimento mostrando a discriminação de correlação. Quatro registros cumulativos de sessões completas de 4 pombos são mostrados. No início da sessão, a correlação entre bicar e alimentar-se é positiva; se uma bicada ocorresse em qualquer parte do intervalo de tempo programado, a comida era entregue no final do intervalo. Seguindo a primeira linha vertical, a correlação mudou para negativa; uma bicada durante o intervalo cancelava a apresentação de comida no final do intervalo. A taxa do bicar caiu. Seguindo a segunda linha vertical, a correlação reverteu-se para positiva, e a taxa de bicar voltou a subir. Adaptado de Baum (1981a).
Um teste
Para testar o ponto de vista molar de contingência ou correlação, podemos aplicá-la para explicar um enigma. W. K. Estes (1943; 1948) relatou dois experimentos onde pré-treinou ratos em duas condições: (a) um tom foi pareado com comida sem nenhuma barra presente, e (b) pressão à barra foi treinada fazendo a comida contingente à pressão à barra. A ordem das duas condições não fez diferença para os resultados. O ponto-chave era que o tom e barra nunca ocorriam em conjunto. Estes testou os efeitos do tom o apresentando enquanto a pressão à barra era colocada em extinção (sem comida). A Figura 14 mostra um resultado típico. A pressão à barra diminui ao longo do intervalo de 5 minutos, mas cada vez que o tom ocorria, a pressão aumentava.
Este achado apresentou um problema para a visão molecular de Estes – isto é, para uma teoria dependente de contigüidade entre eventos discretos. A dificuldade era que, uma vez que o tom e a barra nunca haviam ocorrido juntos, nenhum mecanismo associativo baseado em contigüidade poderia explicar o efeito do tom na pressão à barra. Além disso, a pelota de comida não poderia fazer a mediação entre o tom e a barra, porque a comida nunca tinha precedido a pressão à barra, apenas a seguido. A “explicação” de Estes era criar o que ele chamou de “estado antecipatório condicionado” (CAS). Ele propôs que pareando o tom com comida resultou na eliciação do CAS pelo tom e que o CAS, então, afetou a resposta de pressão à barra durante o teste de extinção. Esta idéia, entretanto, levanta a questão: “Por que o CAS elevaria a pressão à barra?” Ele também nunca havia precedido a pressão à barra. A “explicação” só faz piorar a questão, porque, enquanto o enigma começou com a explicação para o efeito do tom, agora mudou para uma entidade hipotética não observada, o CAS. A confusão ilustra bem como a visão molecular do comportamento leva a entidades hipotéticas e pensamento mágico. O ponto de vista molecular pode explicar o resultado de Estes somente se utilizar-se de mediadores invisíveis (Trapold & Overmier, 1972). (Veja Baum, 2002, para uma discussão mais completa.)
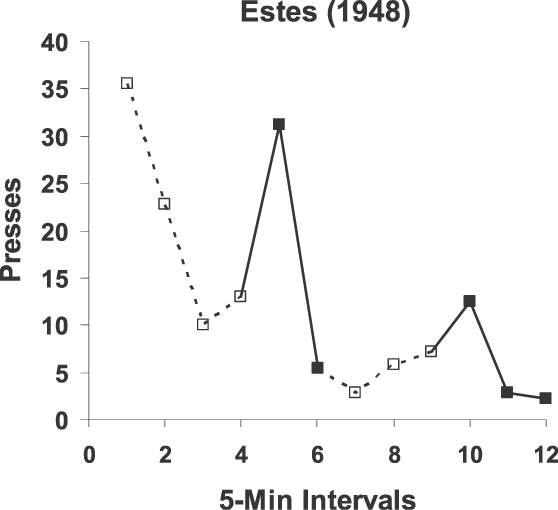
Fig. 14. Resultados de um experimento em que a pressão à barra passando por extinção foi aumentada pela apresentação de um tom que tinha sido previamente pareado com o alimento que tinha sido usado no treinamento de pressão à barra. Porque o tom e a pressão à barra nunca tinham ocorrido juntos antes, o ponto de vista molecular tem dificuldades em explicar o efeito do tom sobre a resposta de pressão à barra, mas o ponto de vista molar explica como a indução da pressão à barra como uma atividade relacionada a alimento. Os dados foram publicados por Estes (1948).
O paradigma molar (por exemplo, Baum, 2002; 2004) oferece uma explicação clara do resultado de Estes. Primeiro, parear o tom com a comida faz do tom um estímulo indutor (um indutor condicional). Isto significa que o tom irá induzir qualquer atividade relacionada à comida. Em segundo lugar, tornar a comida contingente a pressão à barra – isto é, correlacionar pressão à barra a comida – torna a pressão à barra uma atividade relacionada à comida. Assim, quando o tom é reproduzido na presença da barra, induz a pressão à barra. Quando tiramos o foco da contiguidade, o resultado parece óbvio.
Restrição e Conexão
Os efeitos da contingência vão além de simplesmente “fechar o ciclo”. A contingência tem dois efeitos: (a) cria ou causa uma correlação entre a atividade operante e o evento contingente – o equivalente ao que um economista chamaria de uma restrição; (b) como isto ocorre repetidamente, logo faz com que a atividade operante torne-se relacionada ao evento contingente (indutor) – serve para conectar os dois.
A contingência assegura o aumento da atividade operante, mas também restringe o aumento, restringindo os possíveis resultados (alocações). A Figura 15 ilustra estes efeitos. O eixo vertical mostra a frequência de um evento indutor representado como tempo gasto em uma atividade de melhoria de aptidão, como comer ou beber. O eixo horizontal indica o tempo gasto em uma atividade induzida ou operante, como a pressão à barra ou correr na roda. A Figura 15 diagrama situações estudadas por Premack (1971) e por Allison, Miller, e Wozny (1979); assemelham-se também a um diagrama de Staddon (1983; Figura 7.2). Em uma condição de linha de base, sem contingência, muito tempo é gasto na atividade indutora e relativamente pouco tempo é gasto na atividade que irá tornar-se um operante. O ponto rotulado “linha de base” indica esta alocação. Quando uma contingência é introduzida, representada pela linha sólida na diagonal, que indica uma contingência tal como num esquema de razão, uma dada duração da atividade operante permite uma dada duração da atividade indutora (ou certa quantidade de PIE). A contingência restringe as possíveis alocações entre o tempo na atividade contingente (comer ou beber) e o tempo na atividade operante (pressão à barra ou correr). Qualquer alocação que ocorra deve estar sobre a linha; alocações fora da linha não são mais possíveis. O ponto na linha ilustra o resultado usual: uma alocação que inclui mais atividade operante do que na linha de base e menos do PIE do que na linha de base. O aumento da atividade operante teria sido chamado de um efeito de reforçamento, no passado, mas aqui aparece como um resultado da combinação de indução com a restrição da linha de contingência (também conhecido como função de feedback; Baum, 1973, 1989, 1992). A diminuição do PIE poderia ter sido chamada de punição de comer ou beber, mas aqui também é um resultado da restrição imposta pela contingência (Premack, 1965).
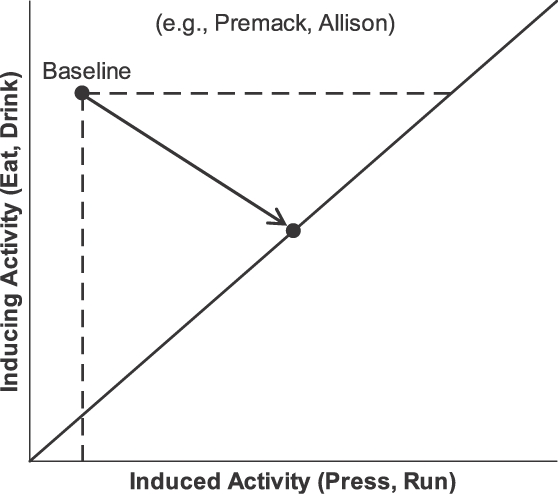
Fig. 15. Efeitos de contingência. Em uma condição de linha de base, sem contingência, pouco da atividade que irá tornar-se um operante (por exemplo, pressão à barra ou correr na roda, a atividade a-ser-induzida) ocorre, enquanto muito da atividade a ser contingente relacionada a PIE (por exemplo, comer ou beber; a atividade de indução) ocorre. Esta é a alocação de linha de base (ponto superior esquerdo e linhas tracejadas). Depois que o PIE (comida ou água) é feito contingente a atividade operante, toda alocação que ocorrer deve estar na linha sólida. A seta aponta para a nova alocação. Tipicamente, a nova alocação inclui um grande aumento na atividade operante sobre a linha de base. Assim, a contingência tem dois efeitos: a) restringir possíveis alocações, e b) tornar o comportamento operante relacionado a PIE de modo que seja induzido em grande quantidade.
Outra forma de expressar os efeitos na Figura 15 pode ser dizer, ao restringir as alocações possíveis, a contingência “agrega valor” à atividade operante induzida. Uma vez que a atividade operante se torna relacionada ao PIE, a atividade operante e o PIE se tornam partes de um “pacote”. Por exemplo, bicar e comer tornam-se partes de uma alocação ou pacote que pode ser chamado “alimentação”, e pode-se dizer que o alimento ou comer emprestam valor para o pacote de alimentação. Se a pressão à barra for necessária para um rato correr em uma roda, a pressão à barra se torna parte da resposta de correr (Belke, 1997; Belke & Belliveau, 2001). Se a pressão à barra for necessária para um camundongo obter material para o ninho, a pressão à barra torna-se parte da construção do ninho (Roper, 1973). A Figura 15 indica que isso não ocorre sem algum custo, também, porque a atividade operante não aumenta o suficiente para levar o nível de comer ao seu nível de linha de base; algum nível da atividade de alimentação é sacrificado em favor de um nível mais baixo da atividade operante. Em um exemplo diário, suponha que Tom e seus amigos assistam futebol todos os domingos e, geralmente, comam pizza enquanto assistem. Fazendo referência à Figura 9, podemos dizer que assistir futebol (E1) é correlacionado com o comer pizza (E2), uma vez que as frequências mais elevadas são as conjunções nas caixas marcadas, e as conjunções nas caixas vazias são relativamente raras. Ao invés de apelar para o reforço ou o fortalecimento, podemos dizer que ver futebol induz a comer pizza ou que as duas atividades formam um pacote com valor mais alto do que qualquer um dos dois isoladamente. Não é preciso introduzir conceito de “valor”, porque tudo que ele significa é que o PIE induz atividades relacionadas ao PIE, mas esta interpretação de “agregar valor” serve para ligar a presente discussão com a economia comportamental. Outra possível ligação seria a de olhar para a demanda como a indução da atividade (por exemplo, trabalho, ou fazer compras) pelo produto demandado e elasticidade como a mudança na indução com a mudança no esquema a de pagamento ou função de feedback. No entanto, um relato quantitativo completo das alocações que realmente ocorrem está além do escopo deste artigo (ver Allison, 1983; Baum, 1981b; Rachlin, Battalio, Kagel, e Green, 1981; e Rachlin, Green, Kagel, e Battalio, 1976 para exemplos). Parte de uma teoria pode ser formulada por meio da quantificação de contingência com o conceito de função de feedback, porque as funções de feedback permitem a especificação mais precisa das variedades de contingência.
Correlações e Funções de Feedback
Toda contingência ou correlação especifica uma função de feedback (Baum, 1973; 1989; 1992). Uma função de feedback descreve a dependência da taxa de resultado (digamos, taxa de disponibilidade de comida) e da atividade operante (digamos, taxa de bicadas). A linha de restrição na Figura 15 especifica uma função de feedback para um esquema de razão no qual a taxa do indutor da contingência é diretamente proporcional à taxa da atividade induzida. A função do feedback para um esquema de intervalo variável é mais complicada, visto que precisa aproximar-se da taxa de resultado programada tal como um assíntota. Veja o exemplo na Figura 16. A curva superior tem três propriedades: (a) ela passa pela origem, pois não existe disponibilidade de comida se não houver atividade operante; (b) se aproxima de uma assíntota; e (c) conforme a atividade operante se aproxima do zero, a inclinação da curva se aproxima à relação recíproca do parâmetro a na equação descrita (1/a; Baum, 1992). Ela restringe possíveis desempenhos, porque qualquer que seja a taxa da atividade operante, a taxa de resultado (indutor) deve estar sobre a curva. A curva inferior sugere uma distribuição de frequência da atividade operante em várias ocasiões. Embora a taxa média de operantes e a taxa média de indutores corresponderiam a um ponto na curva de feedback, a distribuição indica que a atividade operante deveria variar em torno da moda e não necessariamente se aproximaria da taxa de atividade máxima.
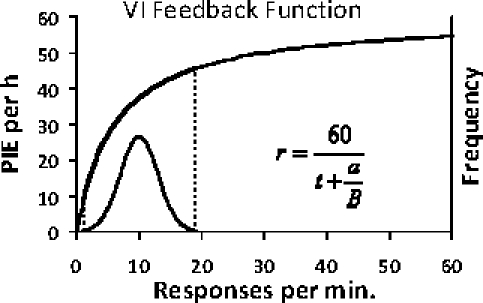
Fig. 16. Exemplo da função de feedback para um esquema de intervalo variável. A curva superior, função de feedback, passa pela origem e se aproxima de uma assíntota (60 PIEs por h; um IV 60s). Sua equação aparece no lado direito inferior. A média do intervalo t é igual à 1.0. O parâmetro a é igual à 6.0. É esperado que a taxa de resposta varie de tempos em tempos, como se vê na distribuição abaixo da função do feedback (frequência está representada no eixo vertical à direita).
A Figura 17 traz dados não publicados obtidos por John Van Sycke (1983), um estudante da Universidade de New Hampshire. Várias sessões de emissão de respostas de um rato em um esquema de intervalo variável foram estudadas através da análise de amostras de intervalos sucessivos de 2 minutos onde foram contados o número de pressões à barra e a disponibilização de comida em cada intervalo. Todos os intervalos de tempo foram agrupados de acordo com o número de pressões à barra. Posteriormente, a taxa de pressão à barra e a taxa de disponibilidade de comida foram calculadas para cada número de pressão. Estes dados são representados pelos pontos que estão traçados no gráfico. A mesma equação como a descrita na Figura 15 se ajusta aos pontos; o parâmetro a estava perto de 1,0. A distribuição de frequência da taxa de pressão é demonstrada pela curva sem símbolos (ordenada direita). Percebe-se que algumas das amostras de 2 minutos não possuem pressões, poucas possuem somente uma (0,5 pressão/minuto), e uma moda aparece em torno de 5 pressões/minuto. Taxas de pressão que superam 15 pressões/minuto (30 pressões em 2 minutos) são raras. Desta maneira, a combinação de indução de pressão a barra através da disponibilidade ocasional de comida junto com a função de feedback resulta em uma performance estável demonstrada pelos resultados.
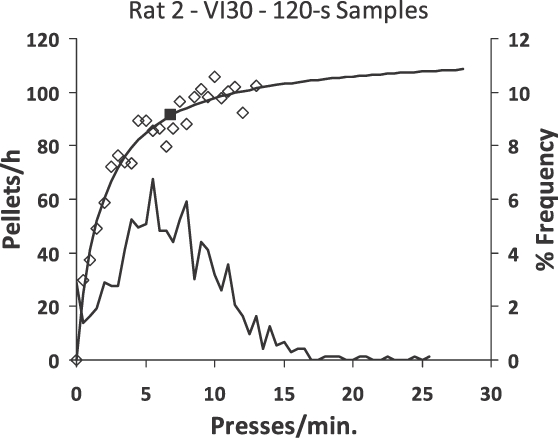
Fig. 17. Exemplo empírico da função de feedback em um esquema de intervalo variável. O número de pressão a barra e número de comida disponibilizada foram avaliados em intervalos consecutivos de 120-s. A taxa de comida e de pressão à barra foi calculada para cada número de pressão por 120 s. Losangos abertos representam a taxa de comida. A equação dd feedback da Figura 15 foi ajustada para estes pontos. A distribuição da frequência (eixo vertical à direita) abaixo mostra a percentagem das frequências de várias taxas de resposta. Os quadrados fechados mostram a média da taxa de resposta.
Embora o parâmetro a esteja perto de 1,0 na Figura 17, ele normalmente excede 1,0, variando em até 10 ou mais quando outros conjuntos de dados estão incluídos na mesma equação (Baum, 1992). Isto representa um quebra-cabeça, pois alguém pode supor que a partir do momento em que a taxa de resposta se reduzir a níveis extremamente baixos, a disponibilidade de comida estaria estabelecida antes de cada resposta, e cada resposta ocasionaria a disponibilização de comida. Isso resultaria na aproximação de taxas semelhantes ao esquema de razão fixa 1 próximo à origem, pela ação da função do feedback. Uma possível explicação da razão pela qual a inclinação (1/a) continua reduzida a 1,0 próxima da origem refere-se ao fato de que quando a disponibilidade de comida torna-se rara, ocorre uma espécie de efeito reestabelecedor, e cada disponibilização de comida induz uma explosão de pressão à barra. Já que a disponibilidade de comida não foi reestabelecida, a explosão normalmente fracassa em produzir comida e desta maneira garante várias pressões para cada vez que comida é disponibilizada.
A relevância da função de feedback tem sido desafiada por dois tipos de experimentos. No primeiro, uma contingência de curto prazo foi comparada a uma contingência de longo prazo. Neste caso, a contingência de curto prazo demonstrou ser capaz de governar o desempenho numa extensão maior do que a contingência de longo prazo (ver, Thomas, 1981; Vangha & Miller, 1984). Nesses experimentos, o bicar a tecla ou a pressão à barra produzia comida na contingência de curto prazo, porém cancelava a comida na contingência de longo prazo. O nível de resposta ficou abaixo do ideal, pois a taxa de comida teria sido mais elevada se nenhuma resposta ocorresse. O fato de que respostas ocorreram nesses experimentos sustenta a presente perspectiva. A comida positivamente correlacionada induz a atividade relacionada à comida (pressão ou bicar); na outra situação onde a comida é negativamente correlacionada, quando ocorre, simplesmente contribui para a indução da atividade operante.
Estes experimentos podem ser comparados a situações como a auto-manutenção negativa (Williams & Williams, 1969) ou o que Hermstein e Loveland (1972) chamou de “esquiva da comida”. Mesmo considerando que bicar a tecla cancela a disponibilidade de comida, ainda assim o pombo bica, pois quando a comida é disponibilizada induz o bicar (incondicionalmente; Figura 1). No entanto, Sanabria, Sitomer e Killen (2006) demonstraram que, sob algumas condições, a contingência negativa é altamente eficaz reduzindo a taxa de bicada a níveis baixos, independentemente do fato da comida estar sendo ocasionada continuamente. Retomando a discussão da Figura 13, a contingência negativa entre bicar a tecla e a comida implica também em uma contingência positiva entre outras atividades e comida, particularmente as atividades de bicar fora da tecla. Considerações similares explicam o contrafreeloading – a observação de que um pombo ou rato irá bicar a tecla ou pressionar a barra mesmo se a mesma comida produzida contingente à bicada na tecla ou pressão à barra estiver livremente disponível (Neuringer, 1969; 1970). A bicada ou pressão são aparentemente induzidas pela comida produzida, ainda que outra fonte de comida esteja disponível. Mesmo no caso em que não exista uma tecla para ser bicada, Palva e Zacny (1980) perceberam que pombos que não haviam passado por treinamento e que eram alimentados em um determinado horário do dia, bicavam por toda parte por volta daquele mesmo horário.
O outro desafio da relevância das funções de feedback é apresentado por experimentos onde a função de feedback é alterada sem nenhuma mudança concomitante no comportamento. Por exemplo, Ettinger, Reid e Staddon (1987) estudaram o efeito da pressão à barra produzindo comida em um esquema onde ambos, tempo e pressão intercambiavam entre si para promover a disponibilidade da comida. Durante taxas de respostas baixas, o esquema se assemelhava a um de intervalo, pois o esquema transcorria principalmente com a passagem do tempo, enquanto que durante altas taxas de respostas, o esquema se assemelhava a um de razão, pois os esquemas transcorriam principalmente devido à emissão de respostas de pressão à barra. Ettinger et al. variaram os esquemas e descobriram que a taxa média de respostas diminuía de um modo linear quando a taxa de comida aumentava, porém esta variação no esquema não teve nenhum efeito nesta relação. Eles concluíram que a função de feedback foi irrelevante. No entanto, os esquemas escolhidos por eles eram todos funcionalmente equivalente a esquemas de razão fixa e os ratos responderam exatamente como responderiam em esquemas de razão-fixa. Em desempenhos de razão fixa, comportamentos induzidos que não sejam o de pressão à barra tendem a limitar-se a períodos imediatamente após o recebimento de comida, tal como demonstrado na Figura 1; uma vez que a pressão à barra inicia-se, existe uma tendência de proceder nessa resposta ininterruptamente até que a comida seja novamente disponibilizada. O período pós-comida aumenta conforme a razão aumenta, conservando a proporcionalidade relativa de pressão à barra e de outras atividades induzidas. A inclinação da função linear de feedback determina o período pós-comida, porém a taxa de resposta em esquemas de razão é indiferente a isso (Baum, 1993; Felton & Lyon, 1966). A redução na média da taxa de resposta observada por Ettinger et al. ocorreu pelo mesmo motivo que acontece com esquemas de razão: o período pós-comida, ainda que menor para razões menores, estava numa proporção crescente do intervalo entre-comida em função da diminuição da razão efetiva. Portanto, a conclusão de que funções de feedback são irrelevantes foi injustificada.
Funções de feedback se tornam indispensáveis quando tentamos entender fenômenos básicos, tal como a diferença entre esquemas de razão e de intervalo. Essas duas funções diferentes de feedback – linear para esquemas de razão (Figura 15) e curvilínea para esquemas de intervalo (Figuras 16 e 17) – explicam a diferença nas taxas de respostas dos dois tipos de esquemas (Baum, 1982b). Em esquemas de razão, o ciclo de contingência demonstrado na Figura 12 inclui somente o feedback positivo e direciona a taxa de resposta em direção ao seu nível máximo frente às restrições da situação. Esse nível máximo de taxa de resposta é necessariamente indiferente à variação da razão e da taxa de comida. Em esquemas de intervalo, onde os níveis de taxa de comida se estabilizam, a comida induz taxas de respostas mais moderadas (Figura 17). Essas diferenças na função de feedback é importante para que se possa compreender essas situações de laboratório e também situações do dia-a-dia que envolvem outras contingências, tais como os esquemas de razão onde o próprio comportamento do indivíduo é responsável pelos resultados, em contraste com contingências envolvendo esquemas de intervalo, nas quais outros fatores fora do controle das pessoas, parcialmente determinam o resultado.
Poder Explicativo
O poder conceitual desse modelo – alocação, indução e contingência – supera aquele do conceito de reforçamento, baseado na contiguidade. Caso este termo seja redefinido e chamado de “reforçamento”, ou receba outro nome (“induzimento”?), ele explicaria uma vasta série de fenômenos. Vimos que este modelo explica resultados padronizados tais como condicionamento operante e respondente, a interação operante-respondente e comportamento adjuntivo. Podemos dizer que é capaz de explicar esquiva e resposta mantida por choque. Explica também o efeito de eventos não-contingentes (ex., “reforçamento não-contingente”, um oximoro). Vamos agora finalizar com alguns exemplos de estudos sobre escolha, estudos sobre controle de estímulos e observações do dia-a-dia.
Dinâmicas da escolha. Em um experimento típico de escolha, um pombo bica em duas teclas de resposta que individualmente operam um distribuidor de comida de maneira ocasional e imprevisível baseado na passagem do tempo (esquema de intervalo variável). Escolha ou preferência é medida como um logaritmo da razão de bicadas em uma chave pelas bicadas na outra. Para olharmos para o reforçamento putativo, nós examinamos o logaritmo da razão de bicada na tecla produtiva (esquerda ou direita) pelas bicadas na outra tecla (direita ou esquerda). Com o procedimento correto, se analisamos a preferência seguida de comida, somos capazes de separar qualquer efeito do fortalecimento do efeito indutor da comida (ex., Cowie, Davison, & Ellife, 2011). Por exemplo, um resultado impressionante demonstrou que comida e estímulos que predizem comida induzem outras atividades que não incluem aquela que produz o resultado (Boutros, Davison, & Elliffe, 2011; Davison & Baum, 2006; 2010; Krageloh, Davison, and Elliffe, 2005).
Para uma total compreensão dos processos de escolha é necessário uma análise de várias escalas de tempo, desde extremamente curtas – focando em relações momentâneas – à extremamente longas – focando em relações estendidas (Baum, 2010). Muitas pesquisas sobre escolha concentraram-se em relações estendidas pelo argrupamento de dados de várias sessões. (Baum, 1979; Herrnstein, 1961). Alguns estudos tem examinado o processo de escolha com foco em situações momentâneas através do agrupamento dos dados de um momento de disponibilização da comida ao outro (Aparicio & Baum, 2006; 2009; Baum & Davison, 2004; Davison & Baum, 2000; Rodewald, Hughes, & Pitts, 2010). Em uma análise mais momentânea se examinaria as mudanças das escolhas nos intervalos entre comida, desde o momento imediatamente após a liberação da comida até a próxima vez em que a comida for disponibilizada. É comum se observar nos resultados dessa análise que imediatamente após a comida haja um impulso a preferir a alternativa que foi efetiva em produzir a liberação da comida (ex., Aparicio & Baum, 2009; Davison & Baum, 2006; 2010). Após um momento inicial de alta preferência, é típico que a escolha tenda à indiferença, apesar de que é possível que nunca se torne realmente indiferente. Esses impulsos de preferência parecem ser um reflexo da indução momentânea de respostas envolvendo somente a alternativa que produz o resultado. Uma análise mais momentânea ainda avaliaria as trocas de preferência entre as alternativas ou as alternações nas aproximações às duas alternativas (Aparicio & Baum, 2006, 2009; Baum & Davison, 2004). Uma aproximação longa à alternativa que produz o resultado acontece segue a liberação da comida produzida por tal alternativa, e isso também parece ser resultado de indução momentânea, assumindo que, por exemplo, bicadas na esquerda associadas com comida induzem bicadas na tecla esquerda enquanto que bicadas na direita associadas com comida induzem bicadas na tecla direita. Em um artigo anterior foi demonstrado que os impulsos de preferência são derivados das aproximações, desta maneira longas aproximações podem ser um reflexo mais direto da indução (Baum, 2010).
Efeito de consequências diferenciais. Em relação ao efeito das consequências diferenciais, uma discriminação é aprimorada quando as diferentes alternativas de respostas produzem resultados diferentes (Urcuioli, 2005). Em um procedimento de tentativa discreta, por exemplo, um estimulo auditivo (S1) é apresentado a ratos em algumas das tentativas e um outro estímulo auditivo (S2) é apresentado em outras tentativas. Na presença de S1, a pressão à barra esquerda (A1) produz comida (X1), enquanto que na presença de S2, a pressão à barra direita (A2) produz um líquido açucarado (X2). A discriminação que se estabelece – pressionar a barra esquerda na presença de S1 e pressionar a barra direita na presença de S2 – é maior nessa situação do que nas condições onde as respostas A1 e A2 produzem a mesma consequência. Esse resultado representa um desafio para o paradigma molecular, pois, focando em contiguidade, o efeito das diferentes consequências pode ser visto como o efeito de eventos futuros sob o comportamento atual. A “solução” encontrada para esta problemática tem sido trazer o futuro para o presente adotando “representações” ou “expectativas” invisíveis geradas por S1 e S2 e supor que esses eventos invisíveis determinem A1 e A2. Na visão molar atual, podemos considerar esse resultado como o efeito da indução diferencial decorrente de consequências diferenciais (Ostlund & Balleine, 2007). A combinação de S1 e X1 faz de S1 um indutor condicional das atividades relacionadas a X1. A contingência entre A1 e X1 faz A1 uma atividade relacionada a X1. Em resultado, S1 induz A1. Da mesma forma, as relações de contingência entre S2 e X2 e as relações de contingência entre A2 e X2, fazem com que S2 induza A2 como uma atividades relacionada a X2. Em comparação, quando A1 e A2 produzem a mesma consequência X, a única base para discriminação é a relação de S1, A1, e X em contraste com a relação entre S2, A2, e X. A1 é induzido através da combinação S1+ X, e A2 é induzido pela combinação S2+X. Já que S1 e S2 são ambos indutores condicionais relacionados a X, S1 tenderia a induzir A2 até certo ponto, e S2 tenderia a induzir A1 até certo ponto, reduzindo portanto a discriminação. Essas relações não ocorrem somente em tentativas isoladas, mas sim como um padrão em muitas tentativas. Já que não existe a necessidade de postular-se eventos invisíveis, essa explicação ultrapassa a explicação molecular com base na plausibilidade e elegância.
Transmissão Cultural. A perspectiva atual oferece uma boa explicação para a transmissão cultural. Analisando a evolução cultural como um processo darwiniano, percebemos que a recorrência de práticas que passam de geração para geração ocorre parcialmente por imitação e parcialmente por regras (Baum, 1995b; 2000; 2005; Boyd & Richerson, 1985; Richerson & Boyd, 2005). Um pombo em um compartimento onde a entrega de comida ocorre ocasionalmente observa outro pombo bicando uma bola de ping-pong e, subsequentemente, bica uma bola similar (Epstein, 1984). A bicada do pombo modelo induz um comportamento similar no pombo observador. Quando uma criança imita seu pai, o comportamento do pai induz o comportamento da criança e, se o comportamento da criança se assemelha ao comportamento do pai, dizemos que a criança imita o pai. Conforme crescemos, outros adultos servem de modelo, particularmente aqueles com sucesso aparente – professores, treinadores, celebridades, e assim por diante – e seus comportamentos induzem comportamentos similares em seus imitadores (Boyd & Richerson, 1985; Richerson & Boyd, 2005). No entanto, um fator crucial para a transmissão são as contingências nas quais o comportamento induzido, fruto de imitação, se envolve. Quando isso acontece, estaria correlacionado com recursos, aprovação, ou oportunidade de acasalamento? Se sim, então agora serão induzidos pelos PIEs e não necessitam mais do modelo. Regras, que são estímulos discriminativos gerados por um falante, induzem o comportamento do ouvinte. O comportamento induzido, se persistir, entra em correlação com ambos os PIEs de curto prazo oferecidos pelo falante e, algumas vezes, com PIEs de longo prazo em relações de longo prazo com o (agora não mais) comportamento governado por regras (Baum, 1995b; 2000; 2005).
Trabalhando por remuneração. Finalmente, vamos considerar um exemplo do cotidiano: o trabalho remunerado. Zack trabalha diariamente como lavador de pratos em um restaurante por 8 horas por dia de terça a sábado. Ao final do seu turno no sábado ele recebe um envelope com dinheiro. Tentar explicar a sua persistência nessa atividade representa uma problemática para o paradigma molecular, pois o dinheiro do sábado é muito distante em tempo do trabalho realizado na terça ou até mesmo de todo trabalho realizado no próprio sábado. Ninguém dá dinheiro a Zack em outras ocasiões – digamos, após cada prato lavado. Para solucionar esse problema, o paradigma molecular tem assumido a existência de reforçadores ocultos que frequentemente fortalecem o comportamento durante a execução do seu trabalho. Ao invés de recorrermos a tais causas invisíveis, podemos adotar a perspectiva molar e afirmar, utilizando o bom senso, que o dinheiro por si só mantém o comportamento de lavar pratos. Nós podemos analisar essa situação da mesma maneira que analisamos o rato que pressiona a barra, porém em uma escala bem maior. O dinheiro induz o lavar os pratos, não somente por uma semana, mas como um padrão de trabalho por muitas semanas. Porém, nós explicamos a primeira semana de Zack no trabalho de outra maneira. Zack traz um padrão de comparecer diariamente no trabalho através do período em que frequentava a escola, o que é induzido pelas promessas de seu chefe. O padrão de trabalho precisa ser também avaliado em um contexto mais amplo envolvendo as atividades de Zack, porque o dinheiro é um meio para atingir uma finalidade, não a finalidade por si só; o dinheiro permite outras atividades, tais como alimentar-se e socializar-se (Baum, 1995a; 2002).
Omissões e inadequações
Como eu mencionei no início, é impossível abordar neste artigo todos os fenômenos conhecidos da análise do comportamento. Este artigo visa estabelecer um modelo conceitual que entrelace esses fenômenos de uma maneira não contemplada pelo modelo tradicional.
Talvez a omissão mais significativa seja a análise completa dos efeitos do contexto ou, o que é tradicionalmente chamado de controle de estímulos. No paradigma molar, o contexto necessita ser considerado na mesma magnitude que o comportamento – estendido no tempo e ocorrendo em diferentes escalas de tempo. Em uma discriminação simples, os estímulos indutores – por exemplo, uma luz vermelha e uma luz verde – podem ser observados em um curto espaço de tempo. No entanto, no pareamento com o modelo, onde um modelo precede a exibição das alternativas de escolha, o período de tempo é maior, pois é necessário incluir o modelo mais a alternativa correta (ou seja, partes). Configurações mais complexas exigem um maior espaço de tempo. Por exemplo, experimentos com contextos relacionais inspirados pela Teoria dos Quadros Relacionais exigem um tempo maior para a especificação de um contexto complexo destinado a uma atividade simples tal como a de uma criança apontando um cartão (Berens & Hayes, 2007). No dia-a-dia, as condições para certos comportamentos podem se estender por semanas, meses ou anos. Um agricultor pode levar vários anos para perceber que a plantação de milho é mais lucrativa que a plantação de alfafa, embora a plantação de alfafa seja mais lucrativa em alguns anos. Eu posso necessitar de muitas interações com uma pessoa durante um longo tempo antes de concluir que essa pessoa é confiável.
Eu gostaria de ter falado mais sobre a metáfora tradicional da força tal como é utilizada para se referir à força de resposta, força de ligação ou força associativa. Uma das virtudes do paradigma molar é que este permite um entendimento sobre o comportamento onde se omite a metáfora da força, substituindo-a por alocação e orientação (Davison & Baum, 2006; 2010). O conceito de extinção de comportamento tem sido estreitamente ligado à noção de força da resposta. O tópico sobre extinção sob a visão do paradigma molar será tratado em um trabalho futuro.
Eu não falei de comportamento verbal. Este tema é abordado no meu livro Compreender o Behaviorismo (Baum, 2005). Este é um tópico importante que deverá ser tratado no futuro, mas pode-se dizer que o paradigma molar observa o comportamento verbal em diversos períodos de tempo. “Enunciação” apresenta-se como um conceito útil, porém existem diversas partes que ocorrem dentro de enunciações semelhantes e que provocam um efeito diferente no ouvinte (ex., “Me deixe ir” versus “Nunca me deixe”) e são induzidas por contextos diferentes. Partes relativamente locais como uma terminação de plural ou um sufixo possessivo são também induzidas por contextos diferentes. Enunciações, no entanto, são partes de atividades mais extensas, tais como conversas. Normalmente, conversas são partes de atividades ainda mais extensas, tal como a construção de um relacionamento ou a tentativa de persuadir alguém a lhe emprestar dinheiro.
Eu falei pouco sobre comportamento governado por regras, o qual se faz muito importante para o entendimento de cultura. Eu discuti esse tópico em artigos anteriores (Baum, 1995b, 2000). O paradigma molar é muito útil para discutir sobre o papel das regras nas relações de curto e longo prazo entre grupos e para indivíduos dentro de grupos.
A lista de omissões é sem dúvida longa, porém o leitor interessado em utilizar esse modelo conceitual irá encontrar muitas maneiras de aplicá-lo.
Conclusões
Juntos, os conceitos de alocação, indução e correlação (ou contingência) sustentam uma série de fenômenos reconhecidos na esfera da análise do comportamento desde os anos de 1950. Eles oferecem uma abordagem mais elegante e plausível do que as velhas noções de reforçamento, fortalecimento, e eliciação. Eles tornam estes conceitos supérfluos.
Indução não resolve somente o problema da origem, como também explica os efeitos dos estímulos PIEs e dos estímulos relacionados a PIEs. A indução explica também os efeitos das contingências, pois uma contingência estabelece uma correlação que converte uma atividade operante em uma atividade relacionada a PIE. Juntas, contingência e indução explicam alocação e escolha. Também explicam as mudanças em alocação – ou seja, mudanças no comportamento. Nenhuma destas considerações necessita do conceito de reforçamento como fortalececimento.
A conhecida formulação do comportamento operante como “Antecedente-Comportamento-Consequência” pode ser conciliada com esta conclusão. Quando “consequências” aumentam uma atividade, elas não a fortalecem, e sim induzem mais da mesma atividade, tal como os “antecedentes” fazem. De fato, a “consequência” é o antecedente do comportamento que se segue, demonstrado nas Figuras 6 e 7. Quando “consequências” reduzem uma atividade, eles não a enfraquecem, mas sim induzem outros comportamentos incompatíveis com tal atividade (Figura 10).
A formulação esboçada aqui parte de uma perspective molar do comportamento que foca no tempo despendido em atividades ao invés de respostas discretas (e.g., Baum, 2002; 2004; Baum & Rachlin, 1969). Criticas à perspectiva molecular baseadas nas respostas discretas e contiguidade temporal como conceitos centrais podem ser encontradas em artigos anteriores (Baum, 1973; 2001; 2002; 2004). Essa discussão evidencia a superioridade da perspectiva molar na tentativa de auxiliar a análise do comportamento a reconsiderar seus conceitos fundamentais e oferecer uma melhor explicação para os fenômenos comportamentais observados no laboratório e na vida diária.
REFERÊNCIAS
Allison, J. (1983). Behavioral economics. New York: Praeger.
Allison, J., Miller, M., & Wozny, M. (1979). Conservation in behavior. Journal of Experimental Psychology: General, 108, 4–34.
Anger, D. (1963). The role of temporal discriminations in the reinforcement of Sidman avoidance behavior. Journal of the Experimental Analysis of Behavior, 6, 477–506.
Aparicio, C. F., & Baum, W. M. (2006). Fix and sample with rats in the dynamics of choice. Journal of the Experimental Analysis of Behavior, 86, 43–63.
Aparicio, C. F., & Baum, W. M. (2009). Dynamics of choice: Relative rate and amount affect local preference at three different time scales. Journal of the Experimental Analysis of Behavior, 91, 293–317.
Azrin, N. H., & Holz, W. C. (1966). Punishment. In W. K. Honig (Ed.), Operant behavior: Areas of research and application (pp. 380–447). New York: Appleton-Century-Crofts.
Baum, W. M. (1973). The correlation-based law of effect. Journal of the Experimental Analysis of Behavior, 20, 137–153.
Baum, W. M. (1974). On two types of deviation from the matching law: bias and undermatching. Journal of the Experimental Analysis of Behavior, 22, 231–242.
Baum, W. M. (1976). Time-based and count-based measurement of preference. Journal of the Experimental Analysis of Behavior, 26, 27–35.
Baum, W. M. (1979). Matching, undermatching, and overmatching in studies of choice. Journal of the Experimental Analysis of Behavior, 32, 269–281.
Baum, W. M. (1981a). Discrimination of correlation. In M. L. Commons, & J. A. Nevin (Eds.), Quantitative analyses of behavior: Discriminative properties of reinforcement schedules (Vol. I, pp. 247–256). Cambridge, MA: Ballinger.
Baum, W. M. (1981b). Optimization and the matching law as accounts of instrumental behavior. Journal of the Experimental Analysis of Behavior, 36, 387–403.
Baum, W. M. (1988). Selection by consequences is a good idea. Behavioral and Brain Sciences, 11, 447–448.
Baum, W. M. (1989). Quantitative prediction and molar description of the environment. The Behavior Analyst, 12, 167–176.
Baum, W. M. (1992). In search of the feedback function for variable-interval schedules. Journal of the Experimental Analysis of Behavior, 57, 365–375.
Baum, W. M. (1993). Performances on ratio and interval schedules of reinforcement: Data and theory. Journal of the Experimental Analysis of Behavior, 59, 245–264.
Baum, W. M. (1995a). Introduction to molar behavior analysis. Mexican Journal of Behavior Analysis, 21, 7–25.
Baum, W. M. (1995b). Rules, culture, and fitness. The Behavior Analyst, 18, 1–21.
Baum, W. M. (2000). Being concrete about culture and cultural evolution. In N. Thompson, & F. Tonneau (Eds.), Perspectives in Ethology (Vol. 13, pp. 181–212). New York: Kluwer Academic/Plenum.
Baum, W. M. (2001). Molar versus molecular as a paradigm clash. Journal of the Experimental Analysis of Behavior, 75, 338–341.
Baum, W. M. (2002). From molecular to molar: A paradigm shift in behavior analysis. Journal of the Experimental Analysis of Behavior, 78, 95–116.
Baum, W. M. (2004). Molar and molecular views of choice. Behavioural Processes, 66, 349–359.
Baum, W. M. (2005). Understanding behaviorism: Behavior, culture, and evolution (2nd edition). Malden, MA: Blackwell Publishing.
Baum, W. M. (2010). Dynamics of choice: A tutorial. Journal of the Experimental Analysis of Behavior, 94, 161–174.
Baum, W. M., & Davison, M. (2004). Choice in a variable environment: Visit patterns in the dynamics of choice. Journal of the Experimental Analysis of Behavior, 81, 85–127.
Baum, W. M., & Rachlin, H. C. (1969). Choice as time allocation. Journal of the Experimental Analysis of Behavior, 12, 861–874.
Belke, T. W. (1997). Running and responding reinforced by the opportunity to run: Effect of reinforcer duration. Journal of the Experimental Analysis of Behavior, 67, 337–351.
Belke, T. W., & Belliveau, J. (2001). The generalized matching law describes choice on concurrent variableinterval schedules of wheel-running reinforcement. Journal of the Experimental Analysis of Behavior, 75, 299–310.
Berens, N. M., & Hayes, S. C. (2007). Arbitrarily applicable comparative relations: Experimental evidence for a relational operant. Journal of Applied Behavior Analysis, 40, 45–71.
Boutros, N., Davison, M., & Elliffe, D. (2011). Contingent stimuli signal subsequent reinforcer ratios. Journal of the Experimental Analysis of Behavior, 96, 39–61.
Boyd, R., & Richerson, P. J. (1985). Culture and the evolutionary process. Chicago: University of Chicago Press.
Breland, K., & Breland, M. (1961). The misbehavior of organisms. American Psychologist, 16, 681–684.
Bullock, D. H., & Smith, W. C. (1953). An effect of repeated conditioning-extinction upon operant strength. Journal of Experimental Psychology, 46, 349–352.
Cowie, S., Davison, M., & Elliffe, D. (2011). Reinforcement: Food signals the time and location of future food. Journal of the Experimental Analysis of Behavior, 96, 63–86.
Davison, M., & Baum, W. M. (2000). Choice in a variable environment: Every reinforcer counts. Journal of the Experimental Analysis of Behavior, 74, 1–24.
Davison, M., & Baum, W. M. (2006). Do conditional reinforcers count? Journal of the Experimental Analysis of Behavior, 86, 269–283.
Davison, M., & Baum, W. M. (2010). Stimulus effects on local preference: Stimulus–response contingencies, stimulus–food pairing, and stimulus–food correlation. Journal of the Experimental Analysis of Behavior, 93, 45–59.
Davison, M., & McCarthy, D. (1988). The matching law: A research review. Hillsdale, NJ: Erlbaum Associates.
Dawkins, R. (1989). The Selfish Gene (New edition). Oxford: Oxford University Press.
Dinsmoor, J. A. (2001). Stimuli inevitably generated by behavior that avoids electric shock are inherently reinforcing. Journal of the Experimental Analysis of Behavior, 75, 311–333.
Eibl-Eibesfeldt, I. (1975). Ethology: The biology of behavior (2nd edition). New York: Holt, Rinehart, and Winston.
Epstein, R. (1984). Spontaneous and deferred imitation in the pigeon. Behavioural Processes, 9, 347–354.
Estes, W. K. (1943). Discriminative conditioning. I. A discriminative property of conditioned anticipation. Journal of Experimental Psychology, 32, 150–155.
Estes, W. K. (1948). Discriminative conditioning. II. Effects of a Pavlovian conditioned stimulus upon a subsequently established operant response. Journal of Experimental Psychology, 38, 173–177.
Ettinger, R. H., Reid, A. K., & Staddon, J. E. R. (1987). Sensitivity to molar feedback functions: A test of molar optimality theory. Journal of Experimental Psychology: Animal Behavior Processes, 13, 366–375.
Felton, M., & Lyon, D. O. (1966). The post-reinforcement pause. Journal of the Experimental Analysis of Behavior, 9, 131–134.
Ferster, C. B., & Skinner, B. F. (1957). Schedules of reinforcement. New York: Appleton-Century-Crofts.
Gardner, R. A., & Gardner, B. T. (1988). Feedforward versus feedbackward: An ethological alternative to the law of effect. Behavioral and Brain Sciences, 11, 429–493.
Herrnstein, R. J. (1961). Relative and absolute strength of response as a function of frequency of reinforcement. Journal of the Experimental Analysis of Behavior, 4, 267–272.
Herrnstein, R. J. (1969). Method and theory in the study of avoidance. Psychological Review, 76, 49–69.
Herrnstein, R. J. (1970). On the law of effect. Journal of the Experimental Analysis of Behavior, 13, 243–266.
Herrnstein, R. J., & Hineline, P. N. (1966). Negative reinforcement as shock-frequency reduction. Journal of the Experimental Analysis of Behavior, 9, 421–430.
Herrnstein, R. J., & Loveland, D. H. (1972). Foodavoidance in hungry pigeons, and other perplexities. Journal of the Experimental Analysis of Behavior, 18, 369–383.
Hinde, R. A., & Stevenson-Hinde, J. (Eds.). (1973). Constraints on learning. New York: Academic Press.
Hineline, P. N. (1977). Negative reinforcement and avoidance. In W. K. Honig, & J. E. R. Staddon (Eds.), Handbook of operant behavior (pp. 364–414). Englewood Cliffs, NJ: Prentice-Hall.
Hineline, P. N. (1984). Aversive control: A separate domain? Journal of the Experimental Analysis of Behavior, 42, 495–509.
Killeen, P. R. (1994). Mathematical principles of reinforcement. Behavioral and Brain Sciences, 17, 105–171.
Kra¨geloh, C. U., Davison, M., & Elliffe, D. M. (2005). Local preference in concurrent schedules: The effects of reinforcer sequences. Journal of the Experimental Analysis of Behavior, 84, 37–64.
Kuhn, T. S. (1970). The structure of scientific revolutions (2nd edition). Chicago: University of Chicago Press. Malagodi, E. F., Gardner, M. L., Ward, S. E., & Magyar, R.
L. (1981). Responding maintained under intermittent schedules of electric-shock presentation: ‘‘Safety’’ or schedule effects? Journal of the Experimental Analysis of Behavior, 36, 171–190.
Mayr, E. (1970). Populations, species, and evolution. Cambridge, MA: Harvard University Press.
Neuringer, A. J. (1969). Animals respond for food in the presence of free food. Science, 166, 399–401.
Neuringer, A. J. (1970). Many responses per food reward with free food present. Science, 169, 503–504.
Ostlund, S. B., & Balleine, B. W. (2007). Selective reinstatement of instrumental performance depends on the discriminative stimulus properties of the mediating outcome. Learning and Behavior, 35, 43–52.
Palya, W. L., & Zacny, J. P. (1980). Stereotyped adjunctive pecking by caged pigeons. Animal Learning and Behavior, 8, 293–303.
Premack, D. (1963). Rate differential reinforcement in monkey manipulation. Journal of the Experimental Analysis of Behavior, 6, 81–89.
Premack, D. (1965). Reinforcement theory. In D. Levine (Ed.), Nebraska symposium on motivation (pp. 123–180). Lincoln: University of Nebraska Press.
Premack, D. (1971). Catching up with common sense, or two sides of a generalization: reinforcement and punishment. In R. Glaser (Ed.), The nature of reinforcement (pp. 121–150). New York: Academic Press.
Rachlin, H., Battalio, R. C., Kagel, J. H., & Green, L. (1981). Maximization theory in behavioral psychology. Behavioral and Brain Sciences, 4, 371–417.
Rachlin, H., & Burkhard, B. (1978). The temporal triangle: Response substitution in instrumental conditioning. Psychological Review, 85, 22–47.
Rachlin, H., Green, L., Kagel, J. H., & Battalio, R. C. (1976). Economic demand theory and psychological studies of choice. Psychology of Learning and Motivation, 10, 129–154.
Rachlin, H., & Herrnstein, R. J. (1969). Hedonism revisited: On the negative law of effect. In B. Campbell, & R. M. Church (Eds.), Punishment and aversive behavior (pp. 83–109). New York: Appleton-Century-Crofts.
Reid, A. K., Bacha, G., & Moran, C. (1993). The temporal organization of behavior on periodic food schedules. Journal of the Experimental Analysis of Behavior, 59, 1–27.
Reid, R. L. (1958). The role of the reinforcer as a stimulus. British Journal of Psychology, 49, 202–209.
Rescorla, R. A. (1968). Probability of shock in the presence and absence of CS in fear conditioning. Journal of Comparative and Physiological Psychology, 66, 1–5.
Rescorla, R. A. (1988). Pavlovian conditioning: It’s not what you think it is. American Psychologist, 43, 151–160.
Richerson, P. J., & Boyd, R. (2005). Not by genes alone: How culture transformed human evolution. Chicago: University of Chicago Press.
Rodewald, A. M., Hughes, C. E., & Pitts, R. C. (2010). Development and maintenance of choice in a dynamic environment. Journal of the Experimental Analysis of Behavior, 94, 175–195.
Roper, T. J. (1973). Nesting material as a reinforcer for female mice. Animal Behaviour, 21, 733–740.
Roper, T. J. (1978). Diversity and substitutability of adjunctive activities under fixed-interval schedules of food reinforcement. Journal of the Experimental Analysis of Behavior, 30, 83–96.
Sanabria, F., Sitomer,M. T., & Killeen, P. R. (2006). Negative automaintenance omission training is effective. Journal of the Experimental Analysis of Behavior, 86, 1–10.
Segal, E. F. (1972). Induction and the provenance of operants. In R. M. Gilbert, & J. R. Millenson (Eds.), Reinforcement: Behavioral Analyses (pp. 1–34). New York: Academic.
Seligman, M. E. P. (1970). On the generality of the laws of learning. Psychological Review, 77, 406–418.
Shapiro, M. M. (1961). Salivary conditioning in dogs during fixed-interval reinforcement contingent on lever pressing. Journal of the Experimental Analysis of Behavior, 4, 361–364.
Sidman, M. (1953). Two temporal parameters of the maintenance of avoidance behavior by the white rat. Journal of Comparative and Physiological Psychology, 46, 253–261.
Sidman, M. (1966). Avoidance behavior. In W. K. Honig (Ed.), Operant behavior: Areas of research and application (pp. 448–498). New York: Appleton-Century-Crofts.
Skinner, B. F. (1938). Behavior of organisms. New York: Appleton-Century-Crofts.
Skinner, B. F. (1948). ‘‘Superstition’’ in the pigeon. Journal of Experimental Psychology, 38, 168–172.
Skinner, B. F. (1950). Are theories of learning necessary? Psychological Review, 57, 193–216.
Skinner, B. F. (1953). Science and human behavior. New York: Macmillan.
Staddon, J. E. R. (1973). On the notion of cause, with applications to behaviorism. Behaviorism, 1(2), 25–63.
Staddon, J. E. R. (1977). Schedule-induced behavior. In W. K. Honig, & J. E. R. Staddon (Eds.), Handbook of operant behavior (pp. 125–152). Englewood Cliffs, NJ: Prentice-Hall.
Staddon, J. E. R. (1983). Adaptive behavior and learning. Cambridge: Cambridge University Press.
Staddon, J. E. R., & Simmelhag, V. (1971). The ‘‘superstition’’ experiment: A re-examination of its implications for the principles of adaptive behavior. Psychological Review, 78, 3–43.
Thomas, G. (1981). Contiguity, reinforcement rate and the law of effect. Quarterly Journal of Experimental Psychology, 33B, 33–43.
Thorndike, E. L. (2000/1911). Animal intelligence. New Brunswick, NJ: Transaction Publishers.
Timberlake, W., & Allison, J. (1974). Response deprivation: An empirical approach to instrumental performance. Psychological Review, 81, 146–164.
Timberlake, W., & Lucas, G. A. (1989). Behavior systems and learning: From misbehavior to general principles. In S. B. Klein, & R. R. Mowrer (Eds.), Contemporary learning theories: Instrumental conditioning theory and the impact of biological constraints on learning (pp. 237–275). Hillsdale, NJ: Erlbaum.
Trapold, M. A., & Overmier, J. B. (1972). The second learning process in instrumental learning. In A. H. Black, & W. F. Prokasy (Eds.), Classical conditioning. II. Current research and theory (pp. 427–452). New York: Appleton-Century-Crofts.
Urcuioli, P. J. (2005). Behavioral and associative effects of different outcomes in discrimination learning. Learning and Behavior, 33, 1–21.
VanSyckel, J. S. (1983). Variable-interval and variable-ratio feedback functions. Unpublished master’s thesis, University of New Hampshire.
Vaughan, W. J., & Miller, H. L. J. (1984). Optimization versus response-strength accounts of behavior. Journal of the Experimental Analysis of Behavior, 42, 337–348.
Watson, J. B. (1930). Behaviorism. New York: W. W. Norton.
Williams, D. R., & Williams, H. (1969). Automaintenance in the pigeon: Sustained pecking despite contingent nonreinforcement. Journal of the Experimental Analysis of Behavior, 12, 511–520.
Zener, K. (1937). The significance of behavior accompanying conditioned salivary secretion for theories of the conditioned response. The American Journal of Psychology, 50, 384–403.
Recebido: 7 de Janeiro de 2011
Aceite final: 6 de Setembro de 2011
Obrigado a Howard Rachlin, John Staddon e Jesse Dallery pelos comentários que ajudaram muito nas primeiras versões deste artigo.
Endereço para correspondência: William M. Baum, 611 Mason #504, San Francisco, CA 94108 (e-mail: wmbaum@ucdavis.edu; ou wbaum@sbcglobal.net).
Artigo original em inglês. Copyright 2012 by the Society for the Experimental Analysis of Behavior, Inc. All rights reserved.

